

기술동향
생체방어펩타이드의 대량생산 및 실용화에 대한 최근 연구동향
- 등록일2015-03-04
- 조회수7400
- 분류기술동향
-
자료발간일
2015-02-25
-
출처
지능형 바이오시스템 설계 및 합성 연구단
- 원문링크
-
키워드
#생체방어펩타이드#대량생산#생체방어#펩타이드#실용화
- 첨부파일
 생체방어펩타이드의 대량생산 및 실용화에 관한 최근 연구 동향-...
생체방어펩타이드의 대량생산 및 실용화에 관한 최근 연구 동향-...
출처 : 지능형 바이오시스템 설계 및 합성 연구단
생체방어펩타이드의 대량생산 및 실용화에 대한 최근 연구동향

경상대학교 조주현 교수
1. 개요
항생제 내성균의
급격한 증가로 인해 항생제 개발의 필요성이 증대되는데도 불구하고 그동안 신규 항생제 개발은 더디게 이루어져 왔다. 지난 40여 년간 새로 출시된
항생제는 oxazolidinone계열에 속하는 linezolid(자이복스, Pfizer), cyclic lipopeptide계의
daptomycin(큐비신, Cubist pharmaceutical) 등 4종에 불과하다(Song, 2012). 또한 현재 개발이 진행 중인
항생제들도 대부분 기존 항생제와 유사한 작용기작을 가져 지속적인 항생제 내성 문제를 유발할 수밖에 없으며, 비특이적인 항균작용으로 인해 체내
정상세균총(normal flora)을 파괴하여 이차감염을 유발하는 등의 문제를 가진다. 따라서 기존의 항생제와는 다른 작용기작을 가지는 동시에
병원균에만 특이적인 항생작용을 나타낼 수 있는 새로운 패러다임의 감염 치료제의 개발이 절실한 실정이다.
최근 들어 기존 항생제의 문제점을
해결할 수 있는 대안으로 생체방어펩타이드(host defense peptide, HDP)가 주목받고 있다. 생체방어펩타이드는
자연면역체계(innate immune system)의 일원으로 모든 생물체가 가지고 있는 10-50개의 아미노산으로 구성된 작은 크기의
펩타이드로서 생물체가 병원균에 감염되었을 때 1차 방어물질로 작용한다(Zasloff, 2002). 이들은 주로 양전하(+2~+9)를 띠며,
30% 정도의 소수성 아미노산 잔기를 포함하고 있는데, 이러한 특징으로 인해 생체방어펩타이드는 음전하를 띤 세균 세포막과 접촉하게 되면 양친성
α-나선구조(amphipathic α-helix) 혹은 β-병풍구조(β-sheet)를 형성하여 세포막 속으로 끼어 들어가 세포막의 전위를
변화시키거나 세포막 자체에 구멍을 내어 세포막을 파괴함으로써 세균을 죽이는 것으로 알려져 있다. 따라서 생체방어펩타이드는 특정 수용체를
타겟으로하는 기존의 항생제에 비해 내성 유발가능성이 현저히 낮다. 이와 더불어 생체방어펩타이드는 신속한 작용 기전, 기존 항생제 내성균에 작용,
항 내독성(anti-endotoxicity) 등의 특징을 가지고 있어 차세대 항생제로서 개발이 적합하다(Hancock & Sahl,
2006).
실제로 hLF1-11(lactoferricin 유도체), iseganan(protegrin 유도체),
omiganan(indolicidin 유도체) 등의 생체방어펩타이드를 항생제 대체물질로 개발하려는 시도가 지난 10년간 꾸준히 있어
왔다(표1). 하지만 대다수의 생체방어펩타이드들은 생리적 염농도(physiological salt concentration)에서 불활성화되며,
혈청과 소화관 내에 존재하는 단백질 분해효소에 의해 빠르게 분해되기 때문에 표적 기관으로의 전달/표적 기관에서의 작용이 어려운 문제점이 있다.
또한 무엇보다도 화학합성(solid-phase chemical synthesis)을 통해 생산할 경우 생산 비용(1g 당 100-600$,
Yeung et al., 2011)이 높아져 경제성이 낮아지기 때문에 실제로 제품화된 것은 아직 없는 실정이다. 따라서 생체방어펩타이드의
경제적인 대량 생산법을 확립하기 위하여 많은 연구가 진행되고 있다. 본 논고에서는 생체방어펩타이드의 대량생산 및 실용화에 대한 최근 연구동향을
살펴보고자 한다.
표1. 임상시험 중인 생체방어펩타이드
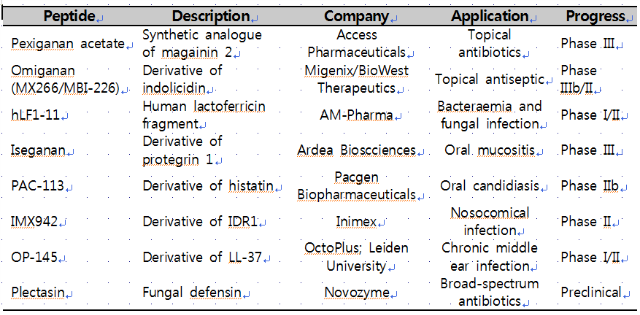
[Modified from Bassetti et al., 2013]
2. 대장균을 이용한 생체방어펩타이드의 생산
현재까지 yeast, insect
cell, mammalian cell, plant 등의 다양한 expression host를 사용하는 non-bacterial
heterologous expression system이 개발되었지만 생체방어펩타이드의 대량생산을 위한 host organism으로는
Escherichia coli가 가장 널리 사용되고 있다(Yin et al., 2007). 생체방어펩타이드를 E. coli에서 발현시킬 경우
생체방어펩타이드의 특징(anti-bacterial activity, small size, high cationic property)으로 인해
숙주 세포에 대해 세포독성을 나타내거나 단백질 분해효소에 의해 쉽게 분해가 되는 문제점이 있다. 따라서 생체방어펩타이드를 carrier
protein과 결합시킨 형태로 발현시킨 뒤, chemical 또는 enzymatic cleavage 과정을 통해 생체방어펩타이드를 얻어내는
fusion expression 전략이 주로 사용되고 있다.
1) Fusion
expression
생체방어펩타이드의 fusion expression에 가장 널리 사용되는 carrier protein은
thioredoxin과 glutathione trasferase(GST)이다(표2). 이들은 재조합 단백질의 E. coli
cytoplasm에서의 solubility를 증가시켜주는 장점을 가지며, 특히 thioredoxin은 size(11.8 kDa)가 작기 때문에
peptide-to-carrier ratio를 증가시켜 높은 수율을 확보할 수 있다. 최근 들어서는 타겟 단백질의 folding과
solubility를 향상시켜주는 것으로 알려져 있는 small ubiquitin-related modifier(SUMO)도 많이 사용되고
있는데, SUMO 역시 thioredoxin과 마찬가지로 size(11.2 kDa)가 작기 때문에 peptide 수율 측면에서 유리하다.
GST는 carrier protein으로서의 기능뿐만 아니라 immobilized glutathione을 이용해 정제가 가능한 affinity
tag으로서의 기능을 동시에 가지는 반면, thioredoxin과 SUMO의 경우 fusion protein을 정제하기 위해서는 추가적인
affinity tag이 필요하며 주로 poly histidine tag이 사용된다.
Thioredoxin과 같은
solubility-enhancing carrier뿐만 아니라, 생체방어펩타이드의 fusion expression에는 inclusion body
형성을 유도하는 PurF fragment, ketosteroid isomerase(KSI), TAF12 histone fold domain 등의
aggregation-promoting carrier도 사용된다(표2). 이러한 aggregation-promoting carrier를 이용한
insoluble expression은 soluble expression에 비해 생체방어펩타이드의 독성 완화와 단백질 분해효소에 의한 분해
방지에 유리한 측면이 있으며, centrifugation을 통해 inclusion body를 쉽게 회수할 수 있는 장점이 있다.
...................(계속)
☞ 자세한 내용은 내용바로가기 또는 첨부파일을 이용하시기 바랍니다.