

기술동향
식물의 생물학적, 비생물학적 스트레스들의 상호작용: 유전자부터 들판까지
- 등록일2015-05-27
- 조회수14861
- 분류기술동향 > 그린바이오 > 농업기술
-
자료발간일
2015-05-26
-
출처
BRIC (생물학연구정보센터)
- 원문링크
-
키워드
#식물#생물학적 스트레스#비생물할적 스트레서 #유전자 #전사체
- 첨부파일
 pdf_0002329.pdf
pdf_0002329.pdf
출처 : BRIC
식물의 생물학적, 비생물학적 스트레스들의 상호작용: 유전자부터 들판까지
저자 : 정수영 (POSTECH)
요약문
식물의 서로 다른 스트레스에 대한 반응들은 매우 복잡하고, 전사체(tranome), 세포, 그리고 생리학적 수준에서의 변화를 수반한다. 최근의 증거들은 다양한 스트레스에 대한 식물의 반응이 정확하게 직면한 환경조건에 관련된 유전자를 발현시키는 프로그램을 활성화시키는 각각의 스트레스에 대한 반응과 다르다는 것을 보여준다. 비생물학적 스트레스는 추가적인 스트레스라기 보다는 생물학적 해충 또는 병원체에 대한 감수성을 감소시키거나 증가시키는 영향을 줄 수도 있다. 이러한 생물학적과 비생물학적 스트레스의 상호작용은 특히 abscisic acid과 같은 서로 증가 시키거나 반대로 작용하는 식물 호르몬 신호전달 경로에 의해서 조율된다. 다양한 스트레스 반응들의 특이성은 복잡한 조절 네트워크 안에서 함께 역할 하는 분자 기작의 영역에 의해서 더 조절 받는다. 전사조절 인자, 인산화 효소, 그리고 활성산소는 heat shock 인자와 small RNA들과 같이 상호작용의 핵심 요소들이다. 이 리뷰는 생물학적, 비생물학적 스트레스 반응의 상호작용을 규명하는데 목적을 두고 있다. 생물, 비생물학적 스트레스 반응 경로를 연결하는 핵심 조절자를 찾는 것이 근본적으로 넓은 범위의 스트레스 내성 작물을 개발하는 기회를 제공할 것이다.
키워드: Abiotic stress, Biotic stress, Hormone signaling, Multiple stress, Stress combination
분야: Agriculture
본 자료는 The
interaction of plant biotic and abiotic stresses: From genes to the field J.
Exp. Bot. 2012; 63(10): 3523-3543 의 논문을 한글로 번역, 요약한
자료입니다.
목차
1. 서론
2. 농업에서의 복합적인 환경스트레스의
도전
3. 조합으로서의 식물 스트레스 연구의 중요성
4. 생물 비생물학적 스트레스의 상호작용에 대한 실험실과 필드에서의
증거
5. 복합적인 스트레스 반응에 대한 전사체 분석
6. 스트레스 결합을 조절하는 호르몬 신호전달 경로
7. 다양한 스트레스
반응에서의 모이는 신호전달 경로
7.1. 전사인자들
7.2. Mitogen-activated protein kinase
(MAPK) cascades
7.3. Heat shock factors (HSFs)
7.4. 활성산소
7.5.
Small RNAs
8. 분자적인 스트레스 결합에서의 특이성 조절
9. 관점들: 새로운 방향에서 식물 스트레스
연구
1.
서론
식물들은 서로 다른 스트레스 요인들이 섞여 있는 환경에 노출된 곳에서 살 수 있도록 진화되었다.
고착생물로서, 식물은 미세한 환경변화의 인지, 복잡한 스트레스 조건에 반응, 그리고 생장과 생식에 필요한 중요한 자원을 유지시키기 위해 피해를
최소한으로 하도록 하는 특정한 기작이 발달되었다. 식물은 다양한 스트레스들의 조합에 직면 했을 때 특이적이고 독특한 스트레스에 대한 반응을
활성화 시킨다. 이러한 측면에서 각각의 스트레스에서 연구된 스트레스 내성 식물의 개발 및 테스트를 위한 현재의 기술은 불충분하다. 특히
생물학적, 비생물학적 스트레스의 조합에서 각각의 신호전달 경로는 반대로 역할을 할 수도 있다. 비록 가치 있을지라도, 그 고립된 스트레스 요소의
연구에 의해 나온 결과들은 식물에게 하나의 스트레스에 대한 영향 이상을 설명할 수 없다. 복합적인 스트레스 반응의 본질을 이해하고, 다양한
스트레스에 내성을 가지면서도 높은 수확량을 유지하는 식물을 개발하기 위한 길을 개척하기 위해 스트레스 연구의 초점을 바꾸는 것은 절실하게
필요하다. 이 리뷰는 분자적 기작이 포함된 설명을 강조하여 식물의 생물학적, 비생물학적 스트레스의 상호작용에 대한 영향을
정리하였다.
2. 농업에서의 복합적인 환경스트레스의
도전
대부분의 작물들은 생장과 생식을 위한 유전적인 잠재력을 온전하게 이룰 수 있는 완벽한 환경에서
자라지는 않는다. 이는 작물의 최대 수확량과 평균 수확량 사이의 차이를 분석함으로 강조되었다. 그 예로, 미국의 연간 밀 수확량 최고 기록은
평균 수확량 보다 8배나 증가 될 수 있다. 수확량의 차이는 스트레스라고 알려진 식물내의 잠재적인 생리적 변화의 손상을 유발하는 좋지 않은
환경조건에 의해서 대부분 설명 될 수 있다. 열, 추위, 가뭄, 염분, 그리고 영양상태 등의 비생물학적 스트레스 요인들은 세계농업에 커다란
영향을 주고 있고, 대부분의 주요작물에 대해 50% 이상의 평균 수익률을 줄인다고 제기 되어 왔다. 또한, 식물들은 곰팡이, 박테리아,
바이러스, 선충, 그리고 초식성 곤충을 포함한 광범위한 해충과 병원균의 공격으로부터 자신을 방어해야만 한다. 각각의 스트레스는 피해를 막고,
생존을 보장하기 위해 식물에 의해 구현된 복잡한 세포, 분자 수준의 반응 시스템을 유발하지만, 종종 생장과 수확량에 손해를
본다.
현재 기후 예측 모델은 평균 글로벌 농업 시스템에 급격하게 영향을 미치는 표면온도가 다음 50~100년 안에 3~5℃ 상승
할 것이라고 나타낸다. 이는 가뭄, 홍수, 그리고 열파의 빈도증가를 수반할 것이다. 특히, 중앙유럽과 중앙아프리카처럼 중간 지역의 따뜻해지고
건조해지는 지역에서는 해수면 상승과 같은 염분의 확장에 의해 농업에 적합한 토지의 감소로 이어질 것이다. 강우량과 온도의 변화는 그 자체로
작물의 영양 품질뿐만 아니라 수율에도 악영향을 미칠 수 있다. 기후 변화는 또한 온도상승에 따른 병원균 확산의 촉진과 같이 해충과 병원균의
서식지 범위에 영향을 미칠 것이다. 따라서 작물은 동시에 발생하는 더 넓은 범위의 환경 스트레스에 맞닥뜨리고 심각한 결과를 가져올 가능성이
있다. 인구증가로 인한 세계 식량 생산성에 대한 증가 압력과 함께 변화하는 기후조건은 스트레스 내성 작물 품종에 대한 요구가 발생한다. 따라서
복합적인 스트레스의 자극에 대한 식물의 반응 기작에 대한 이해는 광범위한 스트레스 내성 작물의 개발을 위한 기회를 제공하는데
중요하다.
3. 조합으로서의 식물 스트레스 연구의
중요성
형질전환과 전통적인 육종을 통한 스트레스 저항성 작물을 만드는 궁극적인 목표는 식물과학의 거의
모든 측면에 녹아 들어 있으며, 공공 및 민간 부문의 연구자들에 의해 추진된다. 많은 분야에서 이미 성공적이다. 예를 들어, 비생물적 스트레스에
대한 저항은 전사인자, 후기 배아발생시 풍부한 (Late embryogenesis abundant) 단백질 및 항산화 단백질 유전자 조작을 통해
다양한 작물에서 입증되었다. 식물 기생 선충에 대한 저항성은 도입 유전자의 발현에 의해 감자, 쌀, 바나나에 부여 되었다. 또한, 생물과
비생물적 스트레스에 대한 내성이 모두 생겼다. 옥수수에서 가뭄 저항성을 유도하기 위한 육종은 기생 잡초 Striga
hermonthica에 추가적인 저항으로 이어졌다. 하지만, 연구 프로그램들은 반드시 다른 생물 또는 비생물학적 스트레스에 대한
감수성을 테스트 하지 않는 특정 스트레스에 대한 내성을 목표로 개발된다. 이는 필드 조건에서 재배 할 때 예상하지 않은 반응을 할 수 있어
예상치 못한 결과를 가져올 수도 있다. 예로서, 카사바 모자이크 바이러스에 대한 저항성을 위해 키워진 다양한 카사바들이 나중에 해충
Bemisia tabaci의 타깃이 되었다. 새로운 가뭄 저항성 벼의 다양체들은 일부 선충의 감염에 민감해졌다, 아마도 뿌리
길이가 길어졌기 때문일 것이다. 그리고, Bacillus thuringiensis (Bt)를 발현시킨 형질전환 목화식물은 높은
온도, 이산화탄소 증가 또는 가뭄 기간 동안 단백질 양이 줄어드는 현상을 보였다.
필드, 실험실 그리고 분자적인 연구들의 늘어나는
양의 증거들은 식물의 특이적으로 조합된 스트레스에 대한 반응은 개별의 스트레스에 대해 연구되어진 내용으로 예측되지 않는 것처럼 추가적인 방법으로
일어나는 것이 아니라는 것을 제시한다. 예를 들어, Mittler의 보고에서 설명되어진 1980년과 2004년 사이의 미국의 전체 농업의 가뭄에
대한 손실은 200억 달러였다. 하지만 가뭄과 폭염의 조합된 스트레스에 대한 손실은 1200억 달러였다. 이는 첫 번째의 유해한 영향을 두 번째
스트레스가 더욱 악화시킬 수 있다는 것을 보여준다. 따라서 야생에서 높은 수확량과 생존력 높은 작물을 개발하기 위해서는 다양한 스트레스 처리에
의한 저항성 형질을 찾기 위한 통합된 접근방법이 필요하다. 식물의 복합적인 스트레스에 대한 반응을 규정하기 위해서, 완전하게 새로운 스트레스의
환경 구성과 동시다발적인 스트레스의 처리가 중요하다.
식물은 구체적이고 복합적인 스트레스 조건에 잘 짜여진 반응을 해야만 한다.
많은 경우 개별의 스트레스는 보통 반대의 반응을 이끌어 낸다. 예를 들어, 높은 온도 스트레스는 보통 식물의 잎의 온도를 낮추기 위해 기공을
열지만, 가뭄의 스트레스 조건에서는 이는 수분의 손실을 유발하는 불이익을 가져온다. 비슷한 예로, 높은 온도에 의해 유발된 호흡양의 증가는
염분과 중금속의 흡수를 증가시키고 이 요소들에 의한 피해가 증가된다. 식물들은 스트레스에 대한 반응은 비용적으로 생장과 수확량에서의 접합성을
감소시키기 때문에 생장과 스트레스에 대한 방어 사이에서 효율적으로 균형있는 자원의 재분배를 해야 한다. 이 비용은 실험적으로 스트레스 반응
유전자의 형질전환 식물체에서 스트레스 내성은 있지만 생장과 수확량의 감소를 가져온다는 예로서 강조된다. 예를 들어, 애기장대의
DREB1A 또는 벼의 DREB1A는 냉동과 건조에 대한 저항성을 주지만 심각한 생장을 유발시킨다. 애기장대에
나무콩의 proline-rich 단백질의 지속적인 발현은 삼투, 염분, 그리고 고온 스트레스에 내성을 가졌지만 크기에서 작게 자란다. 식물의
방어에 대한 비용은 여러 가지의 다른 스트레스 반응에 특정 유전자와 요소가 역할을 한다면 감소할 것이다. 스트레스 반응 경로 사이에서의 결합되는
잠재적 설명이다. 이는 넓은 범위의 스트레스로부터 보호를 위해 축적되는 플로보노이드와 같은 2차 대사물질들, 특정 분자 신호경로, 전사인자,
그리고 생물, 비생물학적 스트레스에 의해 활성화되는 유용단백질을 찾는데 설명된다.
동시다발적으로 발생하는 생물, 비생물학적
스트레스는 복잡한 단계로 나타나며, 다른 호르몬 신호전달 경로를 억제하거나 증가시키는 다양한 호르몬 신호전달 경로에 의해 조절된다. 종종,
식물이 병원균에 노출되면 가뭄과 같은 비생물학적 스트레스 반응이 증가된다, 하지만, 장기간 비생물학적 스트레스는 식물의 방어를 약화시키고 병원균
감염을 증가시킨다. 그럼에도, 생물학, 비생물학적 스트레스의 상호작용에 대한 이해에 집중되어 있는 보고들이 거의 없다. 다음 섹션에서는 생물,
비생물학적 스트레스의 조합에 분자 기작 분석에 의한 스트레스 조절에 대한 연구 내용이 진행될 것이다.
4. 생물 비생물학적 스트레스의 상호작용에 대한 실험실과 필드에서의
증거
복합적으로 발생하는 다른 스트레스 요인들은 추가적이거나 상호관계적일 것이라고 간주 된다. 초기
또는 기존의 스트레스의 존재는 스트레스는 식물의 두 번째 스트레스에 대해 보편적인 스트레스는 축적되는 반응이 있는데 이를 상호작용하는 스트레스
요소라 한다. 두 개의 서로 다른 비생물적 스트레스의 작물에 해로운 상호 작용 효과는 잘 연구되어왔고, 최근에 Mittler와 Blumwald에
의해 리뷰 되었다. 특히 열과 가뭄 스트레스는 각각의 개별 스트레스와 비교하여 작물에 불균형 피해를 유발할 수 있다. 하지만, 생물학적
스트레스가 병원균 또는 병충해와 함께 왔을 때 영향을 조사했을 때, 시간, 자연조건 그리고 스트레스의 강도에 따라 긍정적, 부정적 상호작용이
관찰되었다. 온도의 증가는 박테리아, 바이러스, 곰팡이 그리고 선충의 저항성을 낮추는 부정적 상호영향을 주었고, 밀에서는 6년동안의 실험기간
동안 곰팡이균 Cochliobolus sativus의 민감도가 높은 온도와 상관 관계가 있음이 관찰되었다. 담배와 애기장대에서는
HR 반응과 R 단백질을 매개한 Pseudomonas syringae와 바이러스의 반응이 높은 온도에서는 발휘되지 않아서
병원균의 생장이 증가 되었다. 선충과 진딧물 저항성 유전자인 Mi-1.2 또한 28도 이상에서 비활성화 돠어 감염율이 증가됨이 알려져
있다.
가뭄 스트레스는 식물의 병원균 저항성에 해로운 영향을 유발 할 수 있다. 수수와 콩 모두에서, 가뭄 처리 식물들은 탄저병균
Macrophomina phaseolina에 높은 민감성을 보였다. 유사하게, 가뭄 스트레스는 야자와 포도나무에서 곰팡이균과
박테이라의 잎 타는 현상을 증가 시켰다. 애기장대의 가뭄에 노출은 P. syringae의 높은 감염을
유발시켰다.
반면에 생물학적 스트레스는 병원균 스트레스와 긍정적으로 작용한다. 보리에서, 염분 유도된 삼투압 스트레스의 증가는
흰가루병곰팡이의 저항성과 일치했을 뿐 아니라, 가뭄 스트레스는 곰팡이균 Botrytis cinerea의 저항성을 증가 시킬 수
있다. 병원균은 침임을 위해서 적극적으로 식물의 물과의 관계를 방해 할 수 있다. 예를 들어, 그들은 감염된 조직으로부터 수분의 손실을 줄이기
위해 기공을 닫을 수 있고, 생물학적 스트레스에 대한 저항성에 긍정적인 영향을 줄 수 있다. 파밤나방은 토마토 잎에 생물학적 스트레스의 결과불인
방어 화합물의 함유양을 높여서 가뭄 스트레스를 감소시켰다. 바이러스의 감염은 가뭄으로부터의 적극적인 방어를 제공한다. 바이러스에 감염된 토마토,
사탕무 그리고 벼에서 가뭄 영향은 늦게 나타나며, 비감염된 것 보다 잎에 오랫동안 물이 유지된다. 대사물질 분석은 바이러스 감염에 의해 유도되어
축적된 안토시아닌과 같은 삼투방어물질 과 항산화물질 때문일 것이라 제안했다. 유사한 영향이 유익한 미생물에서도 관찰되었다. Timmusk과
Wagner는 뿌리박테리아인 Paenibacillus polymyxa의 처리는 가뭄과 추가적인 박테리아의 공격에 저항성이 있고
ERD15의 발현이 관련 있다는 것을 발견한 첫 번째 사람들이다. 현재, 많은 종류의 박테리아와 수지상체 근균이 많은 작물 종에서 스트레스
내성을 증가시킨다는 것이 알려져 있다. 이러한 미생물들은 작물의 스트레스 내성을 증가시키는데 유전공학과 육종을 대체할 전략을 제공할
것이다.
토양전염성 병원균은 데이터를 얻는 것과, 샘플링 방법의 어려움으로 스트레스 연구에서 심도 있게 연구되지 않았지만 중요하다.
식물 기생충의 감염은 식물 뿌리에 물 흡수를 심각하게 파괴하여 생물학적 스트레스 영향을 악화시킬 수 있다. 그림 1은 가뭄과 선충의 감염이
동시에 영향을 주는 예시이다. 이 경우, 선충은 가뭄 스트레스의 심각성을 개선시켜주었다. 여러 연구들은 선충과 가뭄스트레스가 식물의 생장과
발달에 주는 영향에 대해 수행되었다. 인도에서 토마토에 뿌리혹 선충의 감염과 광물오염 연구는 선충의 감염이 오염을 더욱 악화시켜 시너지 영향을
주어 뿌리혹병을 증가시켰다고 밝혔다. 동시에 일어나는 생물, 비생물학 스트레스의 영향은 긍정적, 부정적으로 상호 영향을 줄 것이다. 이어지는
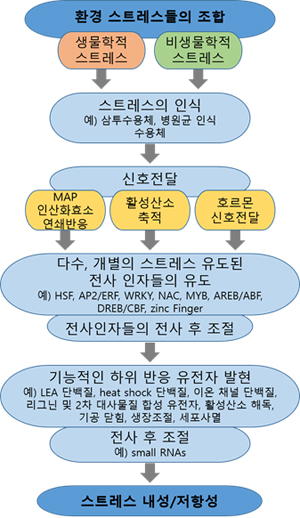
부분에서는 유전자와 이 반응에 관여하는 분자 기작에 대해 집중한다.

그림 1. 생물, 비생물학적 스트레스 조건에서의 벼의 생장
5. 복합적인 스트레스 반응에 대한 전사체
분석
복합적인 스트레스에 대한 식물의 분자적 반응에 대한 조사는 전사의 중첩되는 패턴에 집중되어왔다.
이 영향을 위해 다른 그룹의 식물들에 한 종류 또는 다른 종류의 스트레스를 동시에 처리하고 그 유전자 발현 패턴을 비교하는 여러 종류의 연구들이
수행되었다. 양쪽 모두의 스트레스에 의해 조절되는 중첩되는 세트의 유전자들이 동정되었고, 보편적인 스트레스 반응 또는 신호전달 경로 사이에서
상호 교류하는 포인트가 되는 유전자라고 제안되어 왔다. 이 유전자들이 작물의 스트레스 내성을 증가시키기 위한 타깃으로 추측되어 왔다. 예를 들어
Affymetrix ATH1 마이크로어레이를 이용한 애기장대에 9종류의 다른 생물학적 스트레스에 대한 전사체 비교에서 67개의 유전자들이 각각의
모든 스트레스에서 보편적으로 조절된다는 것이 발견되었다. 이는 각 조건에 반응하는 일반적인 요소로 제안되었다. 사이토크롬 P450 유전자의
발현에 대한 유사한 연구도 다양한 생물학적 스트레스, 병원균 또는 호르몬 처리에 의한 반응에 대해 연구되었다. 생물, 비생물학적 스트레스 모두에
의해 보편적으로 유도된 유전자들이 동정되었다. 따라서 신호경로 사이에 조졸 상호작용이 중요하다. 병아리콩에서 생물, 비생물학적 스트레스에 대한
반응이 758개의 검침 마이크로어레이에 의해 비교되었다.
하지만, 이러한 연구들은 복합적이고 동시다발적인 스트레스 식물 스트레스
반응에 대해 이해하기엔 한계가 있다. 스트레스가 조합적으로 닥쳤을 때 개별의 스트레스에서는 유도되지 않는 특정 유전자들이 활성화된다. 애기장대와
담배 모두에서, 최근 유전체 분석을 통해 식물의 높은 온도와 가뭄이 동시 처리된 스트레스에 대한 분자적 반응은 추가적인 반응이 아니라는 것을
보였다. 애기장대에서 두 개의 스트레스에 의해 특이적으로 조절되는 유전자들 중에는 Heat shock 단백질(HSPs), 단백질
분해효소(proteases), 지질 생합성 효소 그리고 녹말 분해 효소 등이 있다. 뿐만 아니라 MYB 전사인자, 인산화 단백질 그리고 방어에
관련된 단백질들이 산화 스트레스의 보호에 포함되어 있다.
다양한 종에서 추가적인 전사체 연구들은 복합적인 생물, 비생물학적 스트레스
반응을 조절하는 유전자, 호르몬, 그리고 프로세스를 밝혔다. 그리고 스트레스 내성을 증가시키는데 타깃을 제공한다. 해바라기에서 강한 빛과 높은
온도의 조합은 특이적으로 다르게 발현하는 세트의 유전자들을 활성화 시켰다. 밀에서는 가뭄과 고온의 스트레스가 동시에 일어날 때 HSP 발현
패턴을 활성화 시킨다. 생리학적인 수준에서, 가뭄과 고온 스트레스의 도입은 개별적인 스트레스보다 광합성 효율에 굉장한 부정적인 영향을 주었다.
두 개의 기생충 살충제의 처리에 의한 담배에서의 전사체 분석은 각각을 따로 처리한 것과 동시에 처리한 것을 비교할 때 특이적인 유전자 발현
영향을 보였다. 이는 자연에서의 스트레스에 의존적인 유전자 발현을 인지하는 요소 활성요소의 존재를 제공한다. 추가적으로 살충제가 순차적으로
처리되었을 때와 동시에 처리 되었을 때가 다르다. 이는 면역적인 기억의 종류와 같이 생물학적 순차적인 생물학적 공격이 긴 시간의 변화를
전사체적인 것으로 보여주어 시발적인 시스템을 제안한다. 또 다른 두 개의 EST cDNA library를 이용한 곰팡이 균인
Aspergillus parasiticus의 감염과 가뭄에 노출된 땅콩의 분석 연구에서 42개의 유전자는 동시에 처리되었을 때
증가된 반면, 52개의 유전자는 가뭄에서만 증가되었다. 가뭄에 의한 뿌리의 손상은 이 병원균에게 유리하기 때문에, 높은 감염을 위해 곰팡이들이
ABA와 가뭄 신호를 억제할 가능성이 제안되었다.
이러한 종류의 전사체 데이터는 위에서 설명된 다양한 스트레스의 순차적이지 않은
영향에 대한 분자적 설명을 제공한다. 하지만 식물의 동시 처리된 생물, 비생물학적 스트레스 반응에 대한 관심이 증가되고 있는 반면, 그러한
조합된 스트레스에 대한 전체 유전체에 대한 전사체 변화에 대한 보고가 없다.
비록 많은 양의 생산 데이터와 추론 가능한 유용한
통찰력에도, 전사체 연구는 세포 내 단백질의 활성변화에 대한 설명을 제공할 수는 없다. 전사인자와 타깃 유전전자 mRNA 사이의 상관관계에 대한
제안은 적다. 그리고 대부분의 전사인자의 활성 전사 후 조절 또는 인산화를 통하여 조절된다. 추가적으로, 단백질체 데이터와 마이크로어레이를 통해
측정된 전사체의 상관관계는 놀랍도록 낮은 수준이다. 스트레스 신호전달 경로에서 더욱 정확하게 설명되는 구성요소를 얻기 위해, 물질대사, 단백질체
분석의 활용이 증가되고 있다. 전사체에 대한 통합 데이터는 복잡한 식물의 프로세스를 규정하는 대부분의 강력한 장치로서 제공되었다. 추가적으로
호르몬 돌연변이에 대한 연구는 복합적인 스트레스 반응에 대한 방대한 이해를 증가시켰다.
6. 스트레스 결합을 조절하는 호르몬 신호전달
경로
비록 다른 생물학적 공격에 대한 방어는 살리실리산과 자스모닉 산/에틸렌 신호전달경로 사이에서
반대로 나타나는 것과 같이 특화되어 있지만 생물학적 스트레스 반응들은 호르몬인 ABA에 의해 많은 부분 조절 받는다. 하지만, 최근의 발견들은
ABA는 생물학적 신호전달 경로와 상호적으로 그리고 반대로 역할을 한다고 제안하고 있다. 따라서 ABA는 관찰되는 자극된 생물, 비생물학적
스트레스에 반응을 미세하게 조율하는 핵심인 것처럼 보인다. ABA의 처리는 애기장대에서 무병원성 균주인 P. syringae,
토마토에서 B. cinerea와 Erwinia chrysanthemi, 벼에서 아세포 진균인
Magnaportha grisea, 감자에서 병원균인 Phytophthora infestans와
Cladosporium cucumerinum에 대한 민감성이 증가시켰다. 이와 유사하게, ABA의 결여는 높은 수준의 병원균
저항성을 유도 할 수 있다. 그 예로서, ABA에 둔감한 돌연변이인 abi1-1과 abi2-1는 난균류
Peronospora parasitica와 곰팡이 병원균 Fusarium oxysporum에 각각 저항성을
보였고, ABA양이 줄어든 토마토 sitiens 돌연변이는 PR1과 같은 살리실리산 의존적인 발현이 증가될 뿐만 아니라 B. cinerea에
대한 저항성도 증가 되었다. ABA의 처리는 애기장대와 담배에서 살리실리산 유도의 상하경로 모두의 전신획득 저항성 (systemic
acquired resistance, SAR)을 억제할 뿐 아니라, 리그닌과 페닐프로파노이드와 같은 중요한 방어 화합물의 축적도 억제한다.
살리실리산 또한 생물학적 스트레스 신호에 개입한다. 옥수수에 외부에서 처리된 살리실리산은 가뭄에 대한 민감성을 이끌었던 반면 애기장대에서
인위적인 전신획득 저항성의 유도는 비생물학적 스트레스 반응의 억제를 이끌었다. 벼에서 곰팡이균인 M. grisea의 저항성은
정확한 ABA와 SA양 사이에서의 균형에 의해 매개된다. PDF1.2와 같은 방어 유전자의 ABA 매개 억제에 의해 보여지는 것과 같이 ABA는
또한 JA와 에틸렌 방어 신호와 반대로 작용한다. 다음으로 에틸렌 처리는 ABA 신호전달의 부정적인 조절자인 ABI1과 ABI2를 활성화
시킨다. 이러한 결과들에도 불구하고, ABA는 병원균 방어 시스템에 긍정적인 영향을 줄 수도 있다. Melotto는 ABA 유도된 기공 닫힘이
살리실리산 신호전달이 필요한 열려 있는 기공을 통한 식물로의 미생물 침투를 막기 위한 방어 전략임을 찾아냈다. 추가로, ABA는 박테리아 유도된
callose 생산을 막을지라도, 곰팡이류의 병원균에 저항하는 동안 b-aminobutyric acid (BABA)에 의해 유도되는
callose의 침전에 필요하다. 병원균 반응에 있어서 ABA의 다양한 역할에 대한 새로운 모델이 제시되었다. ABA는 병원균 방어 경로를
지배적으로 억제하는 스트레스 반응의 전반적인 조절자로 여겨진다. 따라서 ABA 합성은 어떻게 식물들이 다양한 스트레스에 반응하는지를 결정하기
위한 중요한 요소이다.
...................(계속)
☞ 자세한 내용은 내용바로가기 또는 첨부파일을 이용하시기 바랍니다.
지식
동향