

기술동향
다양한 환경요인과 식물호르몬의 상호작용에 관한 연구동향
- 등록일2015-09-10
- 조회수9862
- 분류기술동향
-
자료발간일
2015-09-08
-
출처
생물학연구정보센터(BRIC)
- 원문링크
-
키워드
#식물호르몬#환경요인#자연환경#식물기작#환경스트레스
- 첨부파일
 pdf_0002384.pdf
pdf_0002384.pdf
출처 : 생물학연구정보센터(BRIC)
다양한 환경요인과 식물호르몬의 상호작용에 관한 연구동향
목차
1. 서론
2. 본론
2.1 생물학적 환경요인과 식물호르몬의 상호작용
2.1.1 유해균 및 병해충
2.1.2 유익균
2.1.3 기생식물
2.2 비생물학적 환경요인과 식물호르몬의 상호작용
2.2.1 가뭄
2.2.2 침수
2.2.3 기타 비생물학적 환경요인
3. 결론
4. 감사의 글
5. 참고문헌
1. 서론
식물을 공부한 사람이라면 한 번쯤 Lincoln Taiz와 Eduardo Zeiger의 저서인 식물생리학 (Plant Physiology)을 접해봤을 것이다. 2014년 하반기, 이 책의 신판이 발간되었다. 눈치 빠른 사람은 알아차렸겠지만 6판이 발간되면서 몇 가지 재미있는 변화가 생겼다. 제목과 저자에도 약간의 변화가 생겼지만, 목차의 변화가 눈에 띈다. 식물생리학에서 항상 큰 부분을 차지하고 있던 식물호르몬 챕터가 보이지 않는다. 5판의 19장 (Auxin)부터 24장 (Brassinosteroids)까지의 여섯 장이 6판에서는 감쪽같이 사라진 것이다. 이러한 챕터구성의 변화는 식물 생리와 발달에 있어 식물호르몬을 바라보는 시각이 어떻게 변하고 있는지를 잘 보여준다. 5판의 19장에 'Auxin transport regulates ?oral bud development and phyllotaxy'라는 세부 단락이 있는데, 6판에서는 동일 내용을 'Auxin, cytokinins, and strigolactones regulate axillary bud outgrowth'라는 제목으로 다루고 있다. 식물호르몬들이 상호작용 (cross-talk)을 하고 있는 것이 밝혀지면서, 어떤 현상을 바라볼 때 조금 더 거시적인 시각으로 바라보려는 시도가 나타나는 것 같다. 목차를 더 살펴보면 다른 재미있는 변화도 눈에 띄는데, 5판에는 없던 'Biotic Interactions' 장이 생겼다는 점이다. 물론 5판에서도 'Assimilation of Mineral Nutrients' 장이나 'Secondary Metabolites and Plant Defense' 장에서 식물과 다른 생명체 간의 상호작용에 대해서 일부 다루기는 하였으나, 6판에서는 이를 하나의 장으로 엮음으로써 식물과 비생물학적 환경뿐 아니라 생물학적 환경과의 상호작용에 대한 관심도 부각되고 있음을 보여준다.
이러한 '교과서'의 집필 방향은 현 식물학계 추세를 반영하는 것 같다. 최근 우리나라뿐 아니라 미국 등 세계적으로 변화하는 환경에서 식량안보가 이슈화 됨에 따라, 연구비 수급에 있어서도 작물연구가 포함되어야 수월하다고 한다. 실제로 식물학계에서는 여러 가지 환경에서의 작물 생산성이 화두가 되고 있다. 그래서 환경과 식물의 상호작용에 있어서도 생산성과 조금 더 밀접한 주제들이 중점적으로 연구되고 있다. 이에 따라 식물호르몬 분야에서도, 생산성에 직결되는 가뭄이나 병해충, 유익균, 영양상태 등의 환경 조건에서 호르몬이 어떻게 작용하는지에 대한 연구가 활발히 진행되고 있으며, 이런 연구동향은 식물생리학의 집필 방향과도 일치하는 것이다. 이러한 동향을 반영하여, 본 동향리포트에서는 생물학적/비생물학적 환경요인을 나열한 후 이와 관련한 여러 식물호르몬의 상호작용에 대한 최신 연구동향을 소개하고자 한다.
본 리포트 작성을 위하여 주로 최근 3년 간의 리뷰논문을 참고하였으며, 리뷰논문 선정은 현재 학계에 있는 지인의 추천 및 미국 국립생물정보센터 (NCBI Pubmed; http://www.ncbi.nlm.nih.gov/pubmed/)에서 아래의 검색조건을 활용하여 진행하였음을 밝힌다.
표 1. 환경요인과 식물호르몬 상호작용 관련 논문 검색을 위한 NCBI 검색식

2. 본론
2.1 생물학적 환경요인과 식물호르몬의 상호작용
식물은 다른 생명체들과 끊임없이 마주하고 있다. 각종 질병을 유발하는 병원균이나 잎을 갉아먹는 병해충, 그리고 영양분을 갈취해가는 기생식물 등의 유해한 생명체도 있지만, 식물이 살아나가는데 필수적인 유익한 생명체들도 존재한다. 이들과 식물과의 상호작용에 대한 연구가 최근 화제가 되고 있으며 이에 대한 동향을 살펴본다.
2.1.1 유해균 및 병해충
유해균과 병해충으로부터 살아남기 위해 식물은 방어 기작을 갖추고 있다. 이 방어 기작을 작동시키기 위해서는 유해균이나 병해충의 공격이 있을 때 이를 식물 스스로가 인지해야 하며, 이런 신호전달과정에 있어 식물호르몬이 중요한 역할을 한다. 대표적인 방어호르몬으로 salicylic acid (SA)와 jasmonic acid (JA)가 잘 알려져 있지만, 에틸렌이나 abscisic acid (ABA), gibberellic acid (GA), 옥신 등 여러 호르몬이 복잡하게 방어 기작에 참여한다. 식물은 종종 성장저해를 감수해가면서까지 방어 기작을 작동시키는데, 여러 가지 호르몬의 양이나 신호강도가 성장보다는 방어 쪽으로 조절되기 때문이다. 지금부터 방어 기작에 관여하는 식물호르몬 신호전달과정을 조금 더 자세히 살펴보기로 한다.
SA는 주로 기생영양병원균 (biotrophic pathogens)에 대항하는 역할을 한다. 식물이 병원균을 인식하면 먼저 SA 생합성이 촉진된다. 병원균 감염 시 Ca2+ 농도가 변화하고 이것이 SA 생합성효소를 작동시키는 요인이 된다. 합성된 SA에 의해 SA 신호전달과정이 유발되는데, 그 중심에는 NON-EXPRESSOR OF PR GENES1 (NPR1)이 있다. 평소에는 NPR1이 서로 간의 이황화 결합을 통해 올리고머를 형성한 상태로 세포질에 존재한다. 일부 소량의 NPR1 단량체는 핵 안으로 들어가기도 하지만, 곧 유비퀴티네이션 (ubiquitinylation)을 거쳐 프로테아좀 (proteasome)에 의해 분해된다. 이는 불필요한 방어 기작 작동을 방지하려는 것이다. 하지만 병원균에 감염되어 SA가 합성되면 이는 세포의 redox state에 변화를 일으킨다. 이에 의해 NPR1의 단량체가 증가하게 되며, 다량의 NPR1 단량체가 핵으로 들어간다. 핵에서 NPR1 단량체는 TGA타입과 같은 전사조절인자들과 결합하여 PR-1 (PHATOGENESIS-RELATED-1)과 같은 타깃 유전자를 발현시킨다 (그림 1). 곧 NPR1은 인산화가 되며, 이어 유비퀴티네이션을 거친 후 분해된다. 이 턴오버 과정은 아마도 새로운 NPR1 단량체에 의한 타깃 유전자의 발현을 미세하게 조절하기 위해 필요할 것이다 [1].
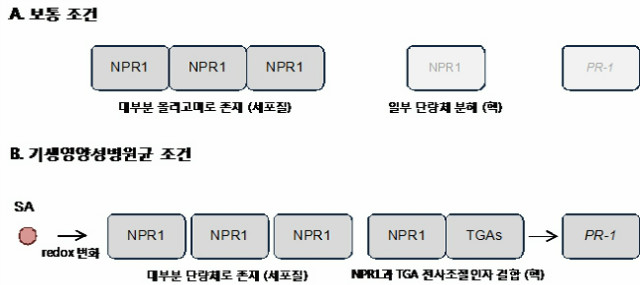
그림 1. 기생영양성병원균 (biotrophic pathogens) 침입에 대응하는 SA 방어 기작
JA는 주로 살생영양성병원균 (necrotrophic pathogens)이나 병해충에 대한 방어 기작에 그 역할을 한다. 이들의 공격이 있으면 JA의 생합성이 촉진되며, 또한 합성된 JA는 JAR1 (JA conjugate synthase)에 의해 높은 활성을 지니는 jasmonoyl-isoleucine (JA-Ile)로 전환된다. 이 활성형 JA-Ile에 의해 JA 신호전달과정이 유발된다. JA 신호전달과정의 중심에는 F-box 단백질인 CORONATINE INSENSITIVE (COI1)와 JASMONATE ZIM (JAZ) 전사억제단백질이 있다. 평소에는 JAZ 단백질들이 JA 신호전달 촉진 전사조절인자인 MYC2, MYC3, MYC4에 결합하여 그 기능을 억제한다. JAZ 단백질의 전사억제 작용을 돕는 조력자들이 있는데, 바로 NOVEL INTERACTOR OF JAZ (NINJA)와 TOPLESS (TPL)이다. NINJA는 JAZ 단백질의 ZIM 도메인과 결합하고, 뒤 이어 TPL을 불러들여 결합한다. NINJA의 ERF-ASSOCIATED AMPHIPHILIC REPRESSION (EAR) 모티프가 TPL을 불러들이는데 그 역할을 한다. 이렇게 형성된 세 단백질의 결합은 평상시에JA 신호를 억제하고 있다. 하지만 병해충 등의 공격으로 JA가 합성되고 JA-Ile의 양이 증가하면, COI1은 JAZ와 함께 JA-Ile 수용체로 작동한다. COI1은 E3 ubiquitin-ligase SKP1-Cullin-F-box compex SCFCOI1의 일부이기 때문에, JA-Ile와 함께 COI1에 결합한 JAZ는 유비퀴티네이션 및 프로테아좀에 의해 분해되어 버린다 (그림 2). JAZ가 분해되면 억제되어 있던 JA 신호전달과정이 살아나게 되며, 이런 JA 반응은 피해받지 않은 식물의 다른 부분에도 영향을 끼쳐 미리 대비하게 한다 [1].
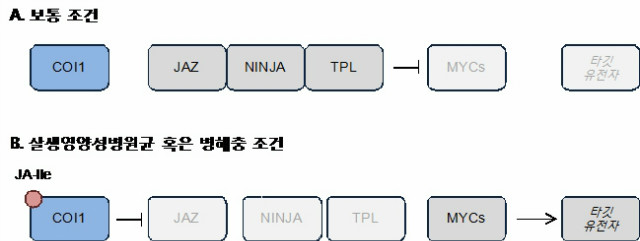
그림 2. 살생영양성병원균 (necrotrophic pathogens)이나 병해충 침입에 대응하는 JA 방어기작
얼핏 보면 SA와 JA는 조력자 같다. 다양한 적으로부터 식물의 방어를 조절하기 때문이다. 실제로 특정 시점이나 농도에서 서로 시너지 효과를 지니기도 하지만, 두 호르몬은 주로 길항작용을 하고 있다고 한다. 이는 애기장대를 포함한 여러 식물에서 밝혀졌다. 이 현상에 대한 이유는 실험실이 아닌 자연에서 찾을 수 있는데, 야생에서의 식물은 다양한 적에 노출되어 있기 때문이다. 앞에서 JA는 주로 살생영양성병원균이나 병해충에, 그리고 SA는 기생영양병원균에 대한 방어 기작에 주로 참여한다고 하였는데, 다양한 적에 대하여 시시각각 효율적으로 대항하기 위해 서로의 기작을 억제하는 방법을 진화시켜 온 것 같다. 이러한 상호 억제 기작 기저에 깔려있는 분자들의 상호작용에 대한 다양한 연구결과 중 일부를 소개하고자 한다. 먼저 앞서 언급한 NPR1이다. 애기장대 npr1 변이체에서는 SA 신호가 억제되었을 뿐만 아니라 SA에 의한 JA 신호억제도 제대로 작동하지 않았다고 한다. NPR1이 SA의 JA 억제작용에 관여하고 있다는 것을 암시하는데, 재미있는 사실은 핵이 아닌 세포질에 위치하는 NPR1만 본 과정에서 역할을 한다고 한다. 애기장대나 벼의 세포질에 NPR1을 과발현 시켰을 때 JA의 신호가 억제되었으며, 벼 세포에서 NPR1을 세포질이 아닌 핵에 과발현 시켰을 때는 이런 억제효과가 보이지 않았다고 한다. NPR1 외에 TGA 전사조절인자도 SA의 JA 억제작용에 관여한다고 한다. 앞에서 TGA는 NPR1 단량체와 결합하여 SA 신호의 타깃 유전자를 발현시킨다고 하였는데, tga256나 tga2356 변이체는 SA 신호가 억제되었음은 물론 SA의 JA 신호억제도 관찰되지 않았다. TGA가 JA타깃 유전자들의 프로모터에 직접 결합하여 억제할 것이라는 가설도 제기되었지만, 여러 연구에 의해 이 가설은 기각되었다. 현재는 TGA가 SA-JA 상호작용에 간접적으로 작용할 것으로 여겨진다. NPR1나 TGA 이외에도 WRKY 전사조절인자나 mitogen-activated protein (MAP) kinase인 MPK4 등도 SA의 JA 억제작용에 관여한다고 알려져 있다 [1].
SA와 JA 이외에도 에틸렌, ABA, 옥신, GA, 사이토카이닌 등이 식물의 방어 기작에 참여하고 있다. 이들은 독립적으로 작용하는 것이 아니라 서로 상호작용을 하며, 이는 주로 SA 및 JA와 연관되어 있다. 대표적인 병원균인 Pseudomonas syringae는 이러한 여러 호르몬의 상호작용을 이용하여 식물의 방어 기작을 회피한다. P. syringae는 식물에게 독소로 작용하는 coronatine (COR)을 분비하는데, 이 COR은 JA-Ile와 분자적 유사성을 지니고 있으므로 COI1와 결합 후 JA 신호를 유발하여 SA 방어 기작을 억제한다. 이외에도 옥신이나 ABA 등의 신호전달 과정을 활용하여 SA의 방어기작을 회피한다고 알려져 있다 (그림 3). 병원균뿐만 아니라 다양한 병해충도 식물호르몬을 조절하여 식물의 방어기작을 회피한다는 연구결과가 많이 알려져 있다 [1].
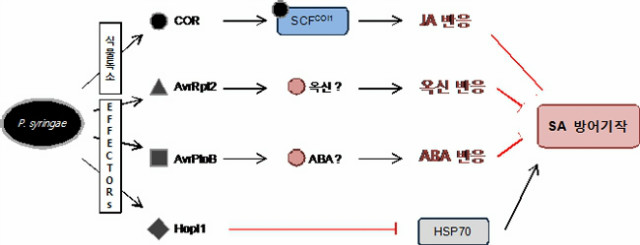
그림 3. 식물의 SA 방어 회피를 위한 Pseudomonas syringae의 식물호르몬 도용 기작
2.1.2 유익균
요즘 여기저기서 '프로바이오틱스'라는 단어가 들린다. 숙주의 건강을 증진시켜주는 미생물을 일컫는데, 인간에게도 질병을 가져다 주는 병원균도 있지만 유산균과 같은 프로바이오틱 세균들은 인간의 건강 유지에 매우 중요하다. 식물에게 있어서도 동물의 유산균 같은 유익균들이 중요한 역할을 하고 있다. 주변에서 나무를 옮겨 심는 작업을 본 적 있다면, 나무 뿌리를 흙으로 감싸서 옮기는 모습을 발견하였을 것이다. 이는 뿌리가 공기 중에 노출되지 않게 하여 수분 증발을 방지하는 데에도 중요한 역할을 하지만, 이미 공생관계를 이루고 있는 수 많은 균들을 함께 옮겨주는 역할도 해준다 (흔히 토양 1그램에는 10억 마리의 세균이 살고 있다고 한다).
식물 유익균은 크게 세 가지로 나누어진다. 균근균 (mycorrhizal fungi), Rhizobium 속 (屬, genus)의 박테리아, 식물성장촉진 근권세균인 PGPR (plant-growth-promoting rhizobacteria)와 PGPF (plant-growth-promoting fungi)가 그것이다. 균근균은 식물뿌리가 닿지 않는 토양으로부터 미네랄 등의 양분을 식물에게 전달해준다. Rhizobium 박테리아는 콩과식물 뿌리에 뿌리혹을 생성하여 공기 중의 질소를 고정하는데 중요한 역할을 한다. PGPR과 PGPF는 식물성장촉진물질을 합성하거나 질병을 억제함으로써 식물의 성장을 돕는다. 이렇게 식물에게 유익한 균들은 앞에서 언급한 식물의 방어기작을 회피해야지만 함께 살아나갈 수 있는데, 이 역시 식물호르몬의 관점에서 풀어보려는 시도가 많이 이루어지고 있다.
PGPF 중 하나인 Piriformospora indica는 SA 방어기작을 억제하기 위해 JA 신호전달과정을 이용한다. P. indica 외에 균근이나 뿌리혹을 통한 공생의 경우에도 초기 접촉 시 숙주 식물의 뿌리에 JA가 일시적으로 축적되는 것을 확인하였는데, 이 역시 SA의 방어기작을 조절하여 공생을 수월하게 하기 위한 기작이라고 볼 수 있다. 많은 PGPR이나 PGPF는 놀랍게도 식물호르몬을 직접 생성하여 방어 기작을 조절하기도 하는데, 옥신이나 GA를 집적 합성하여 SA 방어 기작을 억제하는 것이 그 예이다. 식물호르몬을 직접 합성하는 대신 호르몬 신호전달과정을 조절하는 effector를 활용하기도 하는데, 이러한 effector에는 균근균인 Laccaria bicolor의 MiSSP7이나 Glomus intraradices의 SP7이 있다. MiSSP7은 엔도사이토시스를 통해 식물 세포로 흡수 된 후 핵으로 이동하여 옥신 반응 유전자들의 발현을 촉진한다. SP7은 에틸렌 신호전달과정을 조절하여 식물 면역반응을 억제하는데, 식물세포의 핵으로 이동한 후 방어 관련 전사조절인자인 ERF19에 결합하여 기능을 방해한다. 이러한 여러 가지 방어회피 기작을 통하여 유익균은 식물과의 공생관계를 성공적으로 형성할 수 있는 것이다 [1].
유익균의 기능 중 또 한 가지는 전신획득저항성 (ISR; induced systemic resistance)을 유도하여, 식물의 전체적인 방어능력을 상승시켜 준다는 점이다. PGPR인 Pseudomonas fluorescens WCS417에 의한 ISR 유도가 그 예이다. WCS417은 식물 뿌리로부터 나오는 양분을 이용하여 살아가는데, 뿌리 아포플라스트에 저분자 물질을 방출함으로써 뿌리에서의 면역반응을 억제한다. 뿌리에서 국소적인 면역반응은 억제되지만 곧 식물전체의 면역신호전달 과정을 활성화시켜 식물이 다양한 유해균이나 병해충에 대응할 수 있게 해준다. ISR은 뿌리에서만 발현하는 전사조절인자인 MYB72 및 에틸렌 신호전달에 의해 유발된다. 이렇게 유발된 ISR은 식물 지상부, 특히 잎에서도 역할을 하는데, 이는 전사조절인자인 NPR1과 MYC2가 관여하는 JA/에틸렌 및 ABA 신호전달과정에 의해 조절된다. 특히 ABA는 칼로스 (callose) 형성에 관여하여 식물의 물리적 방어능력을 증가시킨다 [1].
이렇듯 유익균은 식물호르몬 신호전달과정을 자신들에게 유리하게 활용하여, 유익균 스스로는 물론 식물에게도 다양한 이점을 제공하고 있다. 지금까지 소개한 호르몬 이외에도 비교적 최근에 식물호르몬으로 등극한 스트리고락톤 (strigolactone) 또한 유익균과의 상호작용에 중요한 역할을 한다고 알려지고 있다. 스트리고락톤은 원래 기생식물과의 상호작용에 있어 대표적인 식물호르몬으로 알려져 있기 때문에, 다음 장에서 이를 포함한 스트리고락톤의 역할을 간단하게 소개하고자 한다.
2.1.3 기생식물
식물 중에서도 다른 식물에 기생하여 양분을 갈취하는, 소위 '기생잡초'라고도 불리는 기생식물이 있다. Witchweed (Striga spp.)와 broomrape (Orobanche spp. 및 Phelipanche spp.)가 대표적인데, 이들로 인한 작물 피해는 심각한 수준이라고 한다. 이러한 기생식물과 숙주식물과의 상호작용에 있어서 이소프레노이드 (isoprenoid) 화합물이 중요한 역할 (기생식물 뿌리발아유발)을 한다. 이런 역할을 하는 이소프레노이드로는 dihydrosorgoleone, sesquiterpene lactones, 그리고 스트리고락톤 (strigolactone)이 알려져 있고, 이중 스트리고락톤은 최근 식물호르몬으로서의 역할도 한다는 것이 밝혀졌다. 숙주식물은 뿌리에서 근권으로 스트리고락톤을 분비하고, 분비된 스트리고락톤은 굉장히 낮은 농도 (10-7-10-15 M)에서도 기생식물의 뿌리발아를 촉진한다. 기생식물은 며칠 내로 숙주식물에 점착하여 양분을 갈취해야 하며 그렇게 하지 못한 개체는 죽는다 [2].
스트리고락톤은 기생식물 이외에도 유익균인 균근균과 Rhizobium 박테리아와의 공생에서도 중요한 역할을 한다. 먼저 균근균과의 상호작용을 살펴보면, 스트리고락톤은 균근균의 균사분기 (hyphal branching)를 촉진한다고 한다. 10-8 M의 합성 스트리고락톤 (GR24)도 균사분기를 유발한다고 밝혀졌으며, 스트리고락톤 합성이 저해된 완두콩이나 토마토 변이체의 근권에서는 균근류의 형성이 저해되었다고 한다. 이러한 현상을 일으키는 기작을 연구한 결과, 스트리고락톤은 균근균의 유전자 발현을 조절하기에 앞서, 에너지대사를 변화시킨다고 한다. GR24에 노출된 균근균의 균사에서는 1시간 이내에 미토콘드리아의 모양과 밀도, 운동성의 변화가 관찰되었다. GR24에 노출된 Gigaspora rosea 균근균의 균사에서는 노출 후 몇 분 내에 NADH 농도, 탈수소효소 (dehydrogenase) 활성, ATP 양 등 에너지대사 관련 지표가 변하였다. 이들 연구결과는 식물과 균근균의 공생에 있어 스트리고락톤이 중요한 역할을 하고, 스트리고락톤의 균사분기 유도에는 에너지대사 조절이 중요한 역할을 한다는 것을 알려준다. Rhizobium 박테리아과의 상호작용에서도 스트리고락톤이 기능을 한다고 밝혀졌다. Rhizobium 박테리아는 콩과식물 뿌리에서 뿌리혹을 형성하는데, 뿌리혹 형성 과정은 여러 가지 유전자와 펩타이드, 단백질이 관여하는 복잡한 과정이지만 뿌리세포가 뿌리혹으로 분화하는 정확한 분자적 기작은 아직 정확하게 밝혀지지 않고 있다. 스트리고락톤은 식물의 분열조직 (meristem) 발달에 역할을 하기 때문에 이런 뿌리혹 형성과의 관계가 연구되었다. 스트리고락톤 함량이 저하된 완두콩 변이체인 rms1의 연구 결과, 스트리고락톤이 뿌리혹 형성을 촉진한다고 밝혀졌다. 하지만 다른 연구들을 통해 스트리고락톤이 뿌리혹 형성에 필수적이지는 않으며, 다만 최적화를 위해 뿌리혹의 수 등을 조절할 것이라고 제안되고 있다. 스트리고락톤이 뿌리혹 형성에 관여하는 기작은 정확히 밝혀지지 않았지만, 옥신과의 상호작용을 통해 작용할 것이라는 가설이 제기되고 있다 [2].
이러한 스트리고락톤의 역할을 이용하여 아프리카에서는 재미있는 재배법이 이용되고 있다고 한다. 콩과식물과 옥수수를 함께 재배하는 방법인데, 옥수수보다 먼저 자라기 시작하는 콩과식물은 스트리고락톤을 분비하여 근권에 여러 유익균과 공생을 형성한다. 기생식물인 witchweed (Striga spp.) 또한 이 스트리고락톤을 감지하여 발아를 하지만, 안타깝게도 콩과식물은 witchweed의 숙주가 아니다. 곧 이 기생식물은 살아남지 못하게 되고, 잡초가 없는 땅에서 뒤이어 발아한 옥수수는 유익균만 남아있는 좋은 환경에서 성장하게 되는 것이다.
2.2 비생물학적 환경요인과 식물호르몬의 상호작용
식물의 성장과 발달을 조절하는 비생물학적 환경요인으로는 물, 빛, 온도, 영양분 등이 있다. 이러한 다양한 비생물학적 스트레스에 대해 가장 빠르게 생성되는 식물호르몬은 ABA이다. 실제로 가뭄 스트레스가 발생하면, 식물은 잎의 ABA 농도를 최대 50배까지 상승시킨다고 한다. 이는 현재까지 보고된 환경요인에 의한 식물호르몬의 반응 중 가장 극단적인 경우이다. 이번 장에서는 이러한 수분 스트레스에 의한 ABA의 역할을 중심으로, 비생물학적 요인에 대한 식물의 반응과 식물호르몬의 역할을 살펴보기로 한다.
2.2.1 가뭄
수분 부족에 대한 반응은 예전부터 잘 알려져 왔다. 가뭄에 대한 식물의 반응은 주로 기공개폐 조절, 기공 수 조절, 그리고 가뭄 스트레스 유전자 발현으로 나타나고, 이 과정들을 주도하는 식물호르몬은 ABA이다. 기공개폐 조절 기작부터 살펴본다. 가뭄이 오면 식물의 관다발 조직에서 ABA가 축적된 후 공변세포로 이동하고, ABA 신호전달에 의해 기공이 닫혀 증산을 통한 수분손실이 방지된다. 이렇게 가뭄으로부터 기공이 닫히기까지의 분자적 기작을 밝히려는 연구가 많이 이루어져 왔다. 가뭄을 감지한 식물은 특정 조직에서 ABA 합성에 관여하는 유전자인 9-cis-epoxycarotenoid dioxygenase 3 (NCED3)의 발현이 증가되고, ABA는 주로 관다발 조직에 축적된다. ABA는 곧 관다발에서 기공으로 이동하는데, 이때 pH 변화에 의한 수동확산 (passive diffusion)과 수송체에 의한 능동수송을 이용한다. ABA 수송체는 애기장대에서 세 개가 밝혀졌는데, ABC수송체에 속하는 ABCG25와 ABCG40, 그리고 질산 수송체에 속하는 AIT1/NRT1.2/NPF4.6이 그것이다. ABCG25은 ABA를 배출하고, ABCG40은 들여오는 기능을 한다. ABCG25은 관다발에, ABCG40는 공변세포에서 발현하여 관다발로부터 ABA를 배출하여 공변세포로 이동하게 하는데 역할을 할 것이라고 추정되고 있다. 공변세포에서의 ABA는 NADPH oxidase를 통해 활성산소 (ROS; reactive oxygen species)의 생성을 증가시키고, 이렇게 증가된 활성산소는 세포막 칼슘채널을 통한 칼슘이온 유입을 촉진시켜, 공변세포 내 칼슘이온의 양이 증가한다. 이에 의해 S-type (slow-activating)과 R-type (rapid-transient) 음이온채널들이 활성화 되어 공변세포의 음이온이 밖으로 배출된다. 주요 S-type 채널에는 SLOW ANION CHANNEL-ASSOCIATED 1 (SLAC1)이 있으며, SLAC1은 ABA 신호전달과정의 중심에 있는 Snf1-related protein kinase (SRK2E/OST1/SnRK2.6)에 의해 직접 인산화 및 활성화된다. S-type 채널들은 calcium-dependent protein kinase (CDK)인 CDK3, CDK6, CDK21, CDK23에 의해서도 활성화된다는 것이 밝혀졌다. 이러한 음이온 배출에 의해 공변세포의 막은 탈극화 (depolarization) 상태가 되고, 이는 칼륨이온도입 채널인 KAT1/KAT2를 억제하며, 칼륨이온 배출에 관여하는 채널을 활성화시켜 칼륨이온 배출을 촉진한다. 칼륨이온 배출채널로는 GUARD CELL OUTWARD RECTIFYING K+ CHANNEL (GORK) 등이 있다. 이렇게 공변세포 내에는 이온의 양이 줄어들어 삼투압에 의해 물이 빠져나가고, 팽압이 줄어들어 기공이 닫히게 되는 것이다 (그림 4). KAT1도 앞서 언급한 SRK2E에 의해 직접 인산화 된다고 밝혀졌는데, SRK2E에 의해 활성화되는 SLAC1과는 달리 인산화 후 활성이 억제된다고 한다. 최근 KUP/HAK/KT family 칼슘 수송체인 KUP6 또한 가뭄 시 SRK2E에 의해 직접 인산화된다고 밝혀지면서, 식물이 가뭄에 대응하여 기공개폐를 조절하는 기작은 매우 복잡하고, 이 복잡한 과정에서 ABA의 역할이 중요하다는 것이 계속 뒷받침 되고 있다 [3, 4].
...................(계속)
☞ 자세한 내용은 내용바로가기 또는 첨부파일을 이용하시기 바랍니다.