

기술동향
환경 스트레스를 견뎌내는 작물의 유전적 기작
- 등록일2015-11-04
- 조회수15277
- 분류기술동향 > 그린바이오 > 농업기술
-
자료발간일
2015-11-03
-
출처
생물학연구정보센터(BRIC)
- 원문링크
-
키워드
#환경 스트레스#스트레스#작물#유전적 기작#유전
- 첨부파일
 pdf_0002418.pdf
pdf_0002418.pdf
출처 : 생물학연구정보센터(BRIC)
환경 스트레스를 견뎌내는 작물의 유전적 기작
진준영 (CJ제일제당 BIO기술연구소)
목차
1. 도입
2. 수분 과잉 (floods) 대응 기작
2.1 침수 (submergence) 저항성
2.2 침수로부터 탈출
2.3 벼 이외의 다른 작물들의 침수에 대한 전략
2.4 독특한 구조형태학적 항원결정기 (A distinct conformational epitope)
3. 가뭄 (drought) 저항성
3.1 우수한 뿌리 형질
3.2 식물호르몬 Abscisic Acid (ABA) 활용
3.3 가뭄저항성을 위한 복합 메커니즘
4. 염분 (salt) 저항성
5. 알루미늄과 붕소 독성 저항성
6. 필수 영양분 활용성 증대
7. 저온저항성
7.1. 온대작물의 전사조절인자를 통한 냉동저항성 (freezing tolerance)
7.2. 비 (非)온대성 작물의 저온저항성
8. 고온저항성
1. 도입
21세기 인류가 직면하고 있는 중대한 과제는 급변하는 기후와 감소하는 경작지 등의 위기 속에서 식량을 얼마나 안정적으로 수급하느냐이다. 실제로 지난 수십 년 간 열악한 환경 속에 작물 생산성은 줄어들고 있다고 한다. 더욱 심각한 것은 앞으로 홍수와 가뭄, 고온, 저온 등의 발생이 더욱 빈번해 질 것이라고 예측된다는 것이다. 이런 요인에 의해 주요 작물인 옥수수나 밀, 벼의 생산성도 향후 더욱 감소할 것이라고 예상되며, 이는21세기의 식량안보를 직접적으로 위협할 것이다. 1960대 이후로 다양한 유전자원 (germplasm)의 개발 등을 통하여 주요 작물들의 생산성을 상승시키는 노력이 이어지고 있으나, 밀집된 재배 조건은 제한된 경작지 내에서 양분과 수분에 대한경합을 초래하여 기후에 의한 피해를 더욱 증가시키고 있다. 그러므로 이러한 환경에 대하여 내성을 지니고 생산성을 유지할 수 있는 작물의 개발은 인류의 숙제인 것이다.
식물은 세포대사로부터 생리와 발달까지 다양한 조절 기작을 통하여 여러 가지 환경에 적응하며 살아가고 있다. 특히 비생물적 스트레스 (abiotic stress)에 대한 기작이 중요한 적응 요인인데, 비생물적 스트레스란 식물세포의 항상성을 방해하여 결국 식물 성장을 저해하는 기후나 토양 조건을 일컫는다. 수분 과잉이나 부족, 독성 이온 (Al3+, Cl-, Cd2+, Fe2+, Na+ 등), 양분 부족 (Fe3+, N, P, S, Zn2+ 등), 고온, 저온, 대기 오존 (O3) 등이 그 예이다. 이러한 스트레스는 일시적인 스트레스 (transient stress; 낮 동안의 고온 등)와 지속적인 스트레스 (chronic stress; 고염분 토양의 Na+ 등)로 나누어지고, 하루 중 혹은 발달과정 중 언제 스트레스를 받느냐에 따라 작물에게 미치는 영향이 달라진다. 어린 영양생장 시기에 받는 스트레스는 세포분열 및 세포성장을 저해하여 식물 성장을 더디게 하지만 작물의 수확량에 큰 영향을 미치지는 않는다. 반면 생식생장 시기의 스트레스는 작물의 수확량에 심각한 영향을 미친다. 비생물적 스트레스는 복합적으로 찾아오기 때문에 작물 생산성을 유지시키기 위해서는 다양한 적응 기작을 증진시키는 것이 필요하다. 비생물적 스트레스에 의한 피해를 최소화 시키는 것이 결국 농업 생산성을 증대시키는 길이다.
식물은 자연선택에 의한 진화의 결과로 순화 (acclimation; 보통 한 세대에 걸쳐 일어나는 비교적 단기간 내의 적응기작)와 적응 (adaptation; 다세대에 걸쳐 일어나는 적응기작) 기작을 갖추고 있으며, 이를 통하여 스트레스에 대해 내성을 지니거나 스트레스를 회피할 수 있다. 비생물적 스트레스 속에서 높은 생산성을 유지할 수 있게 해주는 형질들은 다양한 종 사이에서 비슷하게 나타난다. 예를 들면 뿌리를 깊게 내림으로써 가뭄에 견디려는 기작은 여러 식물 종 사이에서 보존되어 있는 것이다. 이는 단일 조상으로부터 이어진 계통의 반영일 수도 있으며, 수렴진화 (convergence evolution; 전혀 관계가 없던 생물종들이 비슷한 환경에 적응하기 위해서 서로 유사한 형질을 지니는 현상)의 결과일 수도 있다. 이러한 유용한 형질들은 다양한 유전자원 (germplasm) 속에 존재하고 있으며, 농업에서는 이를 우수 품종으로 개량시켜 오고 있다. 하지만 전통적인 육종에서는 이러한 우수 형질이 구체적으로 어떤 유전자나 생물학적인 기작으로부터 기인하는지를 크게 고려하지는 않는다. 최근 양적형질 (QTL; quantitative trail locus) 맵핑을 통하여 이런 유전적 요인을 찾아내려는 시도가 진행되고 있다. 양적형질과 이와 연관된 유전자들에 대한 분자적 이해를 바탕으로, 품종개량에 우량 유전자를 도입시키는 것이 더욱 용이해졌다. 특히 분자마커를 사용하여 육종 시 작물에 목표 유전자가 도입되었는지 추적이 가능하게 되었다. 이렇듯 우수형질에 대한 분자유전학적 접근은 작물 생산성 이슈를 생명공학적으로 해결하는데 있어 큰 도움이 되는 것이다.
전사조절인자와 수송체, 이 두 가지 유전자군이 식물의 스트레스 내성에 주요한 역할을 한다. 이런 유전자들의 염기서열 (조절부위 포함) 변이나 복제수변이 (CNV; number variation), 유전자 중복 (gene duplication) 후 일어나는 유전자의 새로운 기능성 부여 (neofunctionalization) 등에 의하여 유전형질이 다양해진다. 본 리뷰에서는 수분, 온도, 이온 등에 의한 극한 환경조건에 적응을 도와주는 식물 유전형질에 대해 알아보고자 한다. 또한 포장에서 검증된 비생물적 스트레스에 대한 생명공학적 해결방법들도 소개하고자 한다.
2. 수분 과잉 (floods) 대응 기작
2.1 침수 (submergence) 저항성
아시아에서 재배되는 벼 (Oryza sativa L.)는 야생형인 Oryza rufipogon과 Oryza nivara로부터 곡식낱알의 크기가 크고 촘촘히 들어있는 개체를 찾아서 재배하는 과정으로부터 유래되었다. 각 지역의 자연환경에 우수하게 적응해서 재배되어온 지역품종 (landraces)은 각종 유용한 유전형질들의 기원이 된다. 인도의 지역품종인 Dhalputtia (FR13A)이 대표적인 예인데, 이로부터 침수저항성을 부여하는 유명한 유전형질인 SUBMERGENCE1 (SUB1)이 유래되었다. 벼는 침수상황에서 잠시 성장을 멈추게 하는 휴지전략 (quiescence strategy)을 펼치는데, SUB1이 물 속에서의 신장 (elongation) 억제에 중요한 역할을 한다. 이에 의해 FR13A는 2주 이상의 완전한 침수상황에서도 살아남는다 (논에서 번식하는 반왜성 (semi-dwarf) 품종인 Green Revolution은 침수 상황에서 1주 이내에 죽는다고 한다). SUB1에 대한 맵핑을 진행한 결과 염색체 내의 특정 부위를 찾아내게 되었고, 이 부위는 2~3개의 에틸렌 반응 관련 유전자를 찾아내었다. 이 유전자들은 ethylene-response factor (ERF) DNA-binding proteins를 암호화하고 있다. 이 유전자들은 CNV와 neofunctionalization에 의해 침수저항성을 부여하기도 하고 그렇기 않기도 한다. 이를테면, 침수저항성 지역품종들은 SUB1 부위의 세 개의 ERF (SUB1A, SUB1B, SUB1C) 중 SUB1A-1만 강하게 발현되고 있음을 알 수 있었다. 이 유전자를 침수저항성이 없는 품종인 O. sativa ssp. japonica에 과발현 시켰을 때 침수저항성이 부여된 결과를 통해 SUB1A-1이 침수 저항성에 있어 핵심 유전자임을 다시 한 번 확인하게 되었다.
기체 형태로 존재하는 식물호르몬인 에틸렌은 벼가 물에 잠기게 되면 식물세포 내에 머물게 되고, 이는 SUB1A-1의 발현을 증가시킨다. SUB1A-1는 지베렐린 (GA) 반응을 저해하는 전사조절 인자인 SLENDER RICE 1 (SLR1)과 SLR1-LIKE1 (SLRL1)의 축적을 돕는다. GA는 식물세포의 신장을 촉진하는 호르몬으로 잘 알려져 있는데, 이런 상황에서는 세포가 신장하지 않고 에너지를 보존하게 되는 것이다. 이러한 전략을 통하여 침수상황이 지나고 난 후 ROS로부터의 피해와 공기와 다시 접촉함으로써 생기는 탈수현상으로부터 피해를 최소화하는 것이다. 잎 조직의 손상은 피할 수 없지만, 곁눈 분열조직 (tiller meristem)의 보호를 통하여 침수 이후에도 발달을 재개할 수 있다.
이러한 SUB1A-1이 제공하는 침수저항성을 기존의 생산성이 우수한 벼 품종에서 도입하기 위해 아시아와 아프리카에서는 육종이 활발하게 진행되었다. 이런 지역에서는 홍수 상황이 빈번하게 발생하기 때문이다. 육종의 결과 홍수가 없는 일반 상황에서는 우수 생산성이 그대로 유지되고, 즉, SUB1A-1의 도입이 다른 우수한 형질을 방해하지 않으면서, 침수 상황에서 손실을 최소화하는 우수한 품종들이 개발되었다. 현재 약 1,000만 명의 농업종사자가 Sub1 품종들을 확보하고 있다고 한다.
2.2. 침수로부터 탈출
4미터에 이르는 침수상황에서 오히려 줄기의 신장을 늘려 물 밖으로 성장해버리는 전략도 있다. 이때는 에너지를 세포신장에 투자하여 물보다 높게 자라는 것이다. 벼줄기의 다공성 구조가 뿌리의 통기조직 (aerenchyma)와 연결되어 가스 교환이 가능토록 한다. 태국의 C9285로부터 기인한 세 개의 QTL이 알려져 있으며, 이를 O. sativa ssp. japonica에 도입하였을 때 물 속에서 줄기 신장이 일어남을 확인하였다. 이중 하나는 SNORKEL (SK)라는 유전형질인데, SK1과 SK2라는 ERF가 CNV를 보이고 있다. 이번에도 역할을 시작하는 것은 에틸렌이지만, 앞의 상황과는 다르게 GA 반응을 유도하게 된다. 즉, SK 단백질은 SUB1와는 반대로 GA 반응을 유발시켜, 세포신장을 촉진시킨다. 증가한 GA 합성과 GA 반응은 깊은 침수상황의 벼의 줄기 신장을 위한 전분대사를 가능하게 한다. SK는 아시아의 O. rufipogon 과 Oryza glumaepatula에서 발견되는데, 이는 논에서 재배해오는 과정을 거치면서 Oryza가 원래 지니고 있었던 유전형질이 잃었지만, 여전히 깊은 물에서 재배하는 환경의 벼에서는 이런 형질들이 아직 보존되고 있음을 말해준다. 이상에서 설명한 기작들은 그림 1에서 설명하였다.
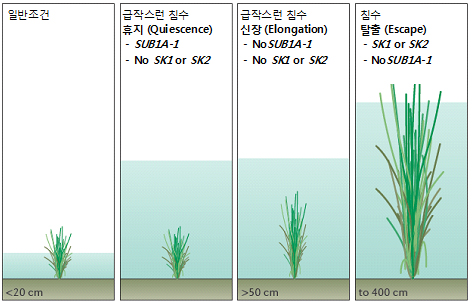
그림 1. 침수에 대한 벼의 저항 기작
2.3. 벼 이외의 다른 작물들의 침수에 대한 전략
벼를 제외한 다른 작물들은 수분 과잉에 대해 굉장히 취약한 모습을 드러낸다. 하지만 애기장대 (Arabidopsis thaliana)에서 발견된 SUB1 및 SK1/2와 유사한 ERF (ERF-VII subclass)가 침수 상황에서 대사적응의 주요한 역할을 하는 것을 발견하게 되었다. 이런 전사조절인자의 축적은 특정 조건에서 일어나며, 산소 접근성에 따라 조절되는 단백질 분해 기작에 관여하고 있음을 알게 되었다. 다양한 종에서 침수나 저산소 조건에서 ERF-VII이 다양한 유전자를 발현시키는데 관여한다는 사실을 이용하여 다양한 작물의 침수저항성을 증가시키는 방법이 고안될 수 있을 것이다.
물에 잠긴 작물의 뿌리는 통기조직의 형성과 뿌리 내부조직의 수베린화 (suberinization)를 통하여, 산소가 뿌리 밖으로 빠져나가는 것을 막는다. 이런 특질에 관여하는 유전형질을 야생형으로부터 재배작물에 성공적으로 도입한 사례가 있다. 바로 Hordeum marinum으로부터 밀, 그리고 Zea nicaraguensis로부터 옥수수에 도입한 사례이다. 이러한 사례들은 야생종으로부터 유용한 침수저항성 형질을 작물에 도입할 수 있는 가능성을 보여준 것이다.
3. 가뭄(drought) 저항성
토양에 수분이 부족하면 토양-뿌리-식물-대기 간의 수분포텐셜 균형이 무너져서, 식물세포에 과삼투압 등의 악영향을 미치게 된다. 가뭄저항성을 증가시키기 위한 육종은 다음 두 가지 요인 때문에 어려운 점이 있다. 첫 번째로, 가뭄 스트레스가 시공간에 따라 너무 다양한 형태로 존재하기 때문에 특정 조건에 부합하는 저항성 요인을 찾아내기 힘들다. 두 번째로, 가뭄은 식물 진화에 있어 매우 중요한 요인이었기 때문에 식물은 이에 적응하기 위한 수 많은 유전형질을 갖추고 있다. 그러므로 하나의 유전자 단독으로 나타내는 효과는 미미한 경우가 많다. 이런 환경과 유전적 다양성이 가뭄저항성 전략을 어렵게 만드는 요인인 것이다. 그럼에도 불구하고, 다양한 조건 안에서도 가뭄저항성을 유효하게 증대시키는 유전형질의 발견을 통하여 이러한 난관을 극복하고 있다.
3.1. 우수한 뿌리 형질
가뭄저항성을 부여하는 유망한 전략 중 하나는 깊은 뿌리 내림을 유도하는 것이다. 이는 수분 이용성을 증대시키는 전략이다. 재배에 널리 이용되는 IR64는 뿌리를 얕게 내리는 벼이다. 뿌리를 깊게 내리는 지역품종인 Kinandang Patong과 IR64의 교배를 통하여 가뭄 상황에서도 생산성을 유지시켜주는 DEEPER ROOTING 1 (DRO1) QTL을 발견하였다. IR64의 DRO1 allele (DRO1-ir)에는 코딩시퀀스에 하나의 염기서열이 빠져있어 불완전한 단백질을 발현하며, 이는 뿌리의 굴중성을 저해한다. 이에 비해 Kinandang Patong allele인 DRO1-kp는 근단(root tip)의 비대칭적 세포신장을 촉진시켜 뿌리가 아래로 뻗게 한다. DRO1-kp를 IR64에 도입하였을 때 뿌리를 깊게 내렸으며, 가뭄상황에서도 높은 생산성을 유지하는 것을 확인할 수 있었다. 이에 더하여 뿌리를 깊게 내리면 수분뿐만 아니라 영양분의 흡수도 증가시켜 가뭄이 아닌 일반 조건에서도 생산성을 증가시키게 된다. 가뭄저항성을 증가시키는 유용한 뿌리형질은 옥수수나 밀, 수수, 보리, 병아리콩 등에서도 알려져 있다.
3.2. 식물호르몬 Abscisic Acid (ABA) 활용
ABA는 식물의 가뭄저항성에 관여하는 대표적인 호르몬이다. 기공의 개폐를 조절하여 증산을 최소화하며, 가뭄저항성에 관여하는 유전자들의 발현을 조절하기도 한다. ABA의 대사와 신호전달, 반응기작 등은 매우 많이 연구가 되었고 잘 알려져 있지만, 이를 실제로 작물의 가뭄저항성 증대에 활용하여 성공한 경우는 극히 드물다. 하지만 ABA 수용체를 명확히 밝혀낸다면, 실제로 적용할 수 있는 여지가 늘어날 것이다. 애기장대에서 ABA가 REGULATORY COMPONENT OF ABA RECEPTOR (RCAR1)와 결합하면, ABA 신호전달과정이 일어난다는 것이 밝혀졌다. 평소에는 type 2C protein phosphatases (PP2Cs)가 ABA 신호전달의 매개체인 SNF-related kinase (SnRK2)를 탈인산화시켜 ABA 반응을 억제하고 있는데, ABA가 RCAR1와 결합하면 이 PP2C의 작용을 억제하는 것이다. RCAR1의 기작이 밝혀졌을 때 즈음에, chemical genomics를 통하여 ABA와 유사한 작용을 하는 화학물질 (small sulphonamide pyrabactin)이 발견되었다. 이 물질은 PYRABACTIN RESISTANCE 1 (PYR1)을 통해서 작용한다는 것도 알려졌다. 이 연구를 통해 13개의 PYR1-Like (PYL) 유전자가 밝혀졌고, RCAR도 이에 속한다. 가뭄 조건에서는 식물 체내에 ABA의 함량이 증가하고, 이는 PYR/PYL-PP2C-SnRK2 모듈을 작동시켜 공변세포의 음이온 채널을 활성화시킨다. 이에 의해 공변세포의 세포막이 탈극화 되며, 최종적으로 기공이 닫히게 된다. 이 모듈은 기공 닫힘을 유도할 뿐만 아니라, 가뭄저항성 유전자의 발현을 증가시키기도 한다. PYR/PYL은 식물 체내에서 발현 위치가 서로 다른 등 특정 조건에서 각자의 역할이 있는 것으로 알려져 있다.
ABA 수용체의 과발현을 통해 가뭄저항성을 증가시킬 수는 있겠으나, 일반 조건에서는 생산성을 감소시킬 우려가 있다. 결국 포장에 성공적으로 적용시키기 위해서는 다양한 ABA 수용체의 발현을 정교하게 조절할 필요가 있는 것이다. 다른 방안으로는 ABA 수용체를 개량하는 것이다. 최근 연구결과에 의하면 애기장대 PYR1의 리간드 결합부위를 개량하여 비제초제 화학물질에 결합하게 만들었고, 이를 이용하여 가뭄저항성을 증가시키는데 성공하였다고 한다. 토마토와 애기장대에 이 개량 PYR1을 발현시킨 후, 잎에 스프레이로 상기 화학물질을 처리했더니 ABA 반응이 일어난 것이다. 이는 ABA를 활용하여 작물의 스트레스에 대한 저항능력을 증가시킬 수 있는 가능성을 단적으로 보여준 사례이다.
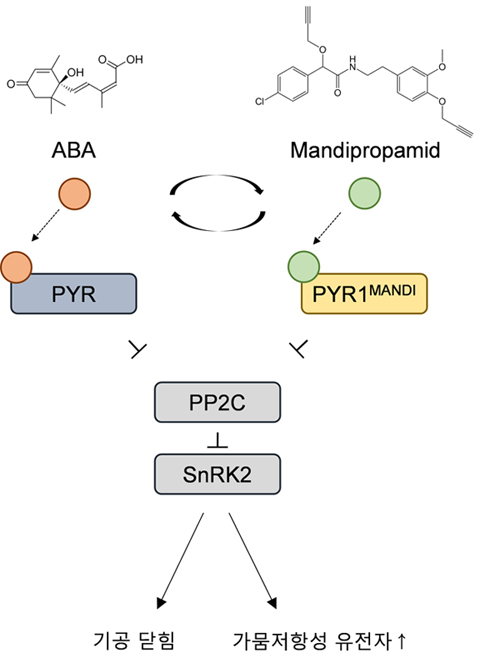
그림 2. ABA 수용체 변이 (PYR1MANDI)와 비제초제 화학물질을 활용한 가뭄저항성 유도
3.3. 가뭄저항성을 위한 복합 메커니즘
앞에서 언급한 뿌리를 깊게 내리는 전략이나 ABA 반응을 이용한 기작 이외에도, 가뭄저항성을 증대시키기 위한 전략이 사용되고 있다. 식물호르몬 사이토카이닌과 에틸렌의 대사조절과 ROS 개량을 통해 stay-green 표현형 (곡물 낱알이 차는 시점에도 엽록소 분해 억제를 통하여 노화가 지연되는 표현형)에 관여하는 유전형질은 곡물육종에 있어 중요한 형질이다. 실제로 다양한 종에 이 형질을 발현시켰을 때 가뭄저항성이 증가하는 것을 확인하였다.
또 다른 연구들을 통하여 신호전달과 조절에 관여하는 다양한 유전자들이 가뭄저항성에 관여한다는 것을 밝혀냈다. 예를 들어 미생물의 RNA결합단백질을 옥수수에 발현시켰을 때 가뭄 상황에서 생산성이 유지되는 것을 확인하였다. 이 외에도 다양한 전사조절인자나 수송체 또한 가뭄저항성에 관여하고 있다.
4. 염분 (salt) 저항성
최근 염분에 의한 농작물의 피해 사례가 증가하고 있다. 현재 전세계 농업생산량의 20%가 염분에 의해 영향을 받고 있다고 한다. 염분 토양은 작물 잎세포 내의 Na+와 Cl- 이온의 농도를 증가시켜 작물생산성을 감소시킨다. 또한 세포성장과 광합성 능력도 저하시키고, 결국 세포독소로 작용하여 잎의 노화를 촉진시켜 탄소동화와 생식기관 형성을 방해한다. 그러므로 뿌리와 지상부의 세포 및 액포로의 Na+ 이온 유입을 조절하여, K+/Na+ 항상성을 조절하고 액포의 삼투조절을 통해 팽압을 유지하는 것은 작물의 염분저항성을 증가시키는데 중요한 역할을 한다.
특히 Na+ 수송체가 많이 연구되어 있으며, 실제 포장에 적용되어 왔다. 세포의 Na+ 항상성을 유지하는데 중요한 수송체로 HIGH-AFFINITY K+ TRANSPORTER1 (HKT1), SALT OVERLY SENSITIVE1 (SOS1), Na+ /H+ EXCHANGER (NHX)가 알려져 있다. HKT1과 SOS1은 세포막에서, NHX는 액포막에서 Na+ 수송을 담당한다. 특히 최근에는 HKT1이 곡물의 염분저항성과 관련하여 부상하고 있다. HKT1;5는 뿌리의 물관으로부터 유조직 (parenchyma) 세포로 Na+를 나르는 역할을 한다. 결국 뿌리 물관의 Na+ 농도가 낮아져서 지상부로의 수분이동을 최소화시키는 것이다. 상업적으로 재배되는 품종인 듀럼밀 (durum; Triticum turgidum ssp. durum)은 HKT1;5이 없어 잎에 Na+이 과도하게 축적되어 염분에 취약한 특성을 드러낸다. 야생에서 밀과 친척뻘인 Triticum monococcum의 유전형질인 Nax2 (TmHKT1;5-A 포함 부위)를 듀럼밀에 도입시킨 결과, 뿌리 물관으로부터 Na+ 배출이 용이해져 잎의 Na+ 농도가 줄어들었으며, 염분 토양에서 생산성이 25% 증가한 것을 확인하였다. 세계 각 지역의 우수 품종들이 염분저항성 획득을 위해 Nax2를 이용한 것으로 알려져 있다. 이 외에도 TmHKT1;4-A를 포함하는 Nax1은 물관으로부터 잎집 (leaf sheath)으로 Na+를 나르는데 참여한다고 밝혀졌다 (그림 2).
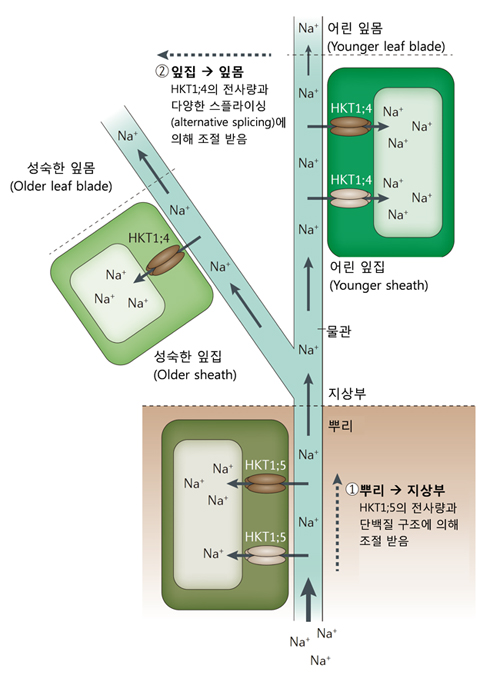
그림 3. 작물의 염분저항성에 기여하는 HKT1 수송체의 역할
어린 시기의 벼에서도 OsHKT1;5이 염분저항성에 관여한다. OsHKT1;5 allele 중 일부는 아미노산 서열에 변이가 있어서 Na+보다 K+를 뿌리 물관 속으로 더 잘 나른다고 한다. 이는 막단백질의 안정성과 인산화 등에 영향을 미쳐 염분저항성에 기여하는 것으로 추정된다. 이 유전자는 어린 벼의 포장에서의 염분저항성으로 잘 알려진 지역품종인 Pokkali의 Saltol QTL 내에 존재하며, 여러 육종 프로그램에 도입되고 있다.
5. 알루미늄과 붕소 독성 저항성
토양에는 Na+ 외에도 식물 생장에 필수적인 Al3+, B, Fe3+, Zn2+ 등이 존재하지만 이들도 과량으로 존재할 때는 식물의 성장에 악영향을 미친다. 또한 Cd2+과 같은 중금속도 존재한다. 그러므로 이들이 식물세포 내에 과량으로 축적되지 않도록 하는 것이 필요하다. 산성 (pH5.5 이하) 토양에서 알루미늄은 대부분 Al3+으로 존재하는데, 이 형태의 이온의 전기화학적 퍼텐셜 (electrochemical potential) 때문에 식물세포막을 쉽게 통과하여 세포 내에 축적된다. 이렇게 축적된 Al3+은 세포의 기본적인 활동을 방해하고 결국 작물의 생산성을 떨어뜨리게 된다. 하지만 다양한 유전자원 CNV 등에 의한 다양한 Al3+저항성 형질이 발견되고 있어, 작물 생산성 보존을 위한 많은 연구가 수행되었다. Al3+에 저항성을 지니는 작물들은 Al3+에 반응하여 일어나는 전사와 전사 후 조절 과정이 강화되어 있음이 확인되었다. 처음으로 알려진 인자로는 세포막에 위치한 유기산 수송체 들이었다. 이들 수송체는 유기산을 식물근권으로 배출하여 주변의 pH를 떨어뜨리고 금속들과 흡착하여 식물이 흡수할 수 없는 형태로 만들어 버린다. 밀의 Al3+-activated malate transporter (TaALMT1)은 유기산 수송체 중에 알루미늄 독성 저항성에 관여한다고 알려진 최초의 수송체이다.
TaALMT1의 기작이 밝혀지고 난 후, 근단에서 구연산을 배출하는 multidrug and toxic compound extrusion (MATE) 수송체들도 일부 곡물에서 Al3+저항성에 관여하는 것이 밝혀졌다. 보리의 HvAACT1 (MATE family)는 피층 (cortical)과 상피층 (epidermal) 세포에서 뿌리 물관으로 구연산을 배출하는데 관여하는데, Al3+저항성이 있는 개체에서는 Al3+에 발현이 더욱 강하게 증가하는 것이 밝혀졌다. 이 개체는 산성토양에서 잘 자라는 개체로 선별되어 왔는데, HvAACT1의 5' untranslated region (UTR)에 1-kb에 달하는 서열의 삽입이 확인되었고, 이에 의해서 근단 피층 및 상피층에서 Al3+에 의한 발현이 더욱 증가하는 것이다. MATE 수송체들의 형질전환에 의하여 Al3+저항성이 증가하는 것은 확인되었지만, 이것이 작물생산성으로 이어졌는지 밝혀지지는 않았다. 벼에서는 유기산 배출에 의한 Al3+ 저항성 부여가 많이 알려지지는 않았다. 대신 세포막의 Al3+ Nramp transporter 1 (NRAT1)이 Al3+ 흡수와 격리를 통해 저항성에 관여하고 있음이 밝혀졌다.
붕소 (B)는 각종 식물에서 분열조직 발달 등에 관여하는 필수 미량원소이지만, 환경에서 부족하거나 과량으로 존재하는 경우가 빈번하다. 최근에 보리와 밀에서 붕소저항성 관련 유전자들이 밝혀졌는데, 붕소가 부족한 환경에서 자라는 품종들은 이 유전자들을 보유하고 있지 않다. 보리의 지역품종인 Sahara 3771에서 강하게 발현되는 유전자인 Bot1 (붕소 배출 수송체 유전자)의 CNV가 붕소저항성과 관련이 깊은 것이 알려졌다. 또한 붕소 흡수에 관여하는 aquaporin-type B transporter (HvNIP2;1)도 보리의 붕소저항성 인자이다. 붕소저항성 보리의 뿌리에서 이 수송체 유전자는 발현이 거의 되지 않는데, 붕소가 과량으로 존재하는 토양에서는 더욱 발현이 감소하여 붕소흡수를 최소화 한다. 밀에서는 A. tauschii 유래의 B transporter-like gene (Bot-B5b)이 붕소저항성에 관여한다고 알려져 있다. 붕소저항성 밀 품종에서는 이 유전자 발현이 강화되어 있으며, 반대로 발현이 감소한 종에서는 붕소 독성에 취약함을 드러내었다. 남부 호주에서 경작되는 많은 우수한 밀 품종들에 Bot-B5가 도입되어 있고, 붕소가 과량으로 존재하는 토양에서 우수한 생산성을 보이고 있다.
6. 필수 영양분 활용성 증대
세계적으로 과열된 작물의 재배는 농경지의 양분은 지속적인 감소를 초래하고 있다. 그렇기 때문에 모델 식물을 통한 필수 양분 (Fe2+, N, P, Zn2+ 등)의 수송체들에 대한 연구가 지속되고 있다. 가뭄저항성에서도 보았듯이 뿌리의 형태는 양분 흡수, 특히 P의 흡수에 있어 중요한 영향을 미친다. 주변의 무기인 (Pi)이 부족해질 때 이를 감지하고 반응하는 기작과 Pi의 흡수능력을 강화시키는 기작은 서로 밀접하게 연관되어 있다. 이를테면 뿌리에서만 발현하는 Pi 수송체 발현 강화, 유기산의 배출, 수지상체 균근 (arbuscular mycorrhizae)과의 공생, 뿌리의 변형 (곁뿌리 및 뿌리털 생성 증가, 뿌리 각의 변화 등) 등이 이에 해당한다. Pi 부족 시 나타나는 체계적인 반응의 중심에는 microRNA도 있다. 이 microRNA는 뿌리로부터 체관을 통하여 잎으로 이동하여 PHO2 mRNA를 분해시키는데 참여한다 (PHO2는 ubiquitin E2 conjugase로 Pi 부족에 대한 반응을 억제하고 있는 단백질이다).
벼의 Phosphorus uptake 1 (Pup1) QTL은 인도와 방글라데시에서 재배되는 aus 지역품종인 Kasalath에서 발견되었다. 이 QTL은 O. sativa ssp. Japonica는 보유하지 않는 염색체 일부분을 포함하는데, 이 부분에 PHOSPHATE STARVATION TOLERANCE1 (PSTOL1) 유전자가 위치하고 있다. Pi가 부족할 때 뿌리와 지상부에서 발현이 약간 증가하는 PSTOL1은 Pi의 흡수를 촉진시키는데 관여한다. PSTOL1은 초기 뿌리 발달을 증가시켜 전체 뿌리의 바이오매스를 늘리는데 기여하는 인산화효소를 암호화하는 유전자이다. O. sativa ssp. Japonica의 Nipponbare에 Pup1을 도입하였을 때 Pi 부족 상황에서도 내성을 지니는 것을 확인하였으며, O. sativa ssp. Japonica의 IR64 및IR74 품종에서는 같은 결과를 포장에서 확인하였다. Pi가 부족한 지역에서 재배되고 있는 많은 품종들에게 Pup1을 도입하려는 시도는 지금도 계속되고 있다.
7. 저온저항성
온대식물은 진화과정을 거치면서 저온에 대한 저항성을 길러왔다. 사멸에 이르지 않을 정도의 적당히 낮은 온도는 식물로 하여금 월동을 할 수 있을 정도의 저온저항성을 가지게 한다. 식물은 세포막 조성의 변화, 친수성 펩타이드 함량 증가, 삼투압조절물질 (compatible osmolyte) 증가, 항산화 작용 강화 등을 통하여 저온에 대해 적응할 수 있다.
7.1. 온대작물의 전사조절인자를 통한 냉동저항성 (freezing tolerance)
보리와 밀 등을 포함하는 외떡잎 온대작물은 저온저항성과 관련 있는 두 개의 전사조절인자 유전자를 지니고 있다. FROST RESISTANCE1 (FR1)라는 염색체 부위는 VERNALIZATION1 (VRN1) 유전자를, 그리고 FR2 부위는 CBF (DREB) 유전자를 지니고 있다. 다양한 계절에 재배할 수 있는 작물개발을 위한 선별적 육종을 통해 이 염색체 부위에 자연변이가 있음을 알게 되었다. 가을에 파종되는 겨울품종은 월동을 위해 저온을 견뎌내야 한다. 이를 춘화 (vernalization)이라고 부르는데, 이때 CBF 전사조절인자가 관여하고 있다. 이런 겨울품종들은 이듬 해 봄에는 식물이 생식생장기로 전환한다. 겨울품종과 달리 봄품종은 보통 춘화 과정을 필요로 하지 않으며 저온에 대한 적응력도 떨어진다. VRN1은 APETALA-LIKE MADS box 전사조절인자 유전자이며, 이 단백질은 저온과 단일 (short day) 조건에서 식물 내에 축적되어 꽃 분열조직의 형성 및 개화를 조절한다. 겨울에 진입하면 겨울밀과 겨울보리는 VRN1 발현 강화를 통해 저온에 대한 내성을 갖춘다. 이 시기에 VRN1의 발현은 미미하다. 하지만 계속되는 저온과 단일에 대한 노출은 VRN1의 전사량을 증가시키고, 이에 의해 식물은 생식생장기에 돌입하여 개화가 일어난다. 이때 CBF의 발현은 감소하며 식물 성장은 촉진된다. 겨울보리는 개화를 거치면서 VRN1의 프로모터와 첫 번째 인트론 부위 뉴클레오솜의 히스톤 H3 메틸화가 점차적으로 일어나고 이로써 VRN1 발현이 억제되어 간다. 이에 반해 봄보리는 그 부위의 변이로 인해 발현억제 기작이 일어나지 않아 VRN1발현을 계속 유지하고 있으며, 이로 인하여 저온에 취약한 형질을 드러낸다.
FR2/CBF 유전자는 일년생 및 다년생 식물의 저온순화 (cold acclimation)과 냉동저항성 (freezing tolerance)에 관여한다. 작물의 FR2 부위에는 여러 CBF 가 복제되어 있는 것을 확인할 수 있다. 보리와 밀의 봄품종에서는 CBF 유전자의 카피수와 춘화 전의 높은 발현량이 냉동저항성에 기여하고 있다. 그러므로 저온적응은 결국 VRN1의 발현조절 기작 및 CBF 의 발현양과 깊은 관련이 있는 것이다.
7.2. 비 (非)온대성 작물의 저온저항성
벼는 영하의 기온에 매우 취약하다. 하지만 벼의 MYBS3 전사조절인자는 어린 시기의 저온저항성과 관련이 있다. MYBS3는 CBF 경로보다 저온에 대해 느리게 반응하기 때문에 CBF와는 독립적인 저온저항성 경로라고 할 수 있다. 이와 더불어 몇몇 QTL이 저온 상황에서 벼의 생산성을 유지해주는 인자라고 알려져 있다.
8. 고온저항성
식물의 생식생장기에 특히 고온에 약하기 때문에, 고온 역시 작물생산성을 저해하는 요인 중 하나이다. 생식생장기에 고온에 노출되면 웅성불임, 자성불임, 화분과 암술머리와의 상호작용 저해 등의 현상이 나타나기 때문이다. 수정 후 야간의 고온은 벼와 같은 작물 종자의 성숙을 저해하고 결국 수확 후 종자의 질을 떨어뜨리는 요인이 된다. 고온 조건에서도 호흡과 광합성을 유지하기 위해서는 여러 가지 기작들이 작동하는데, 고온에 반응하여 재빨리 유전자의 발현을 변화시키는 것, heat shock protein (HSP)의 증가, 세포막의 변화, 세포골격의 변화, 항산화 기작 등이 이에 포함된다. HSP는 단백질이 올바로 접히게 도와주며 단백질 간의 응집을 막아주기 때문에 고온저항성에 기여한다. 하지만 HSP 발현이 높은 상태로 유지되면, 일반 조건에서 식물의 성장이 저해되는 문제점이 있다.
고온 지역의 유전자원에서 고온저항성에 관련된 형질이 발견되고 있다. 이런 형질을 선별하기 위한 육종이 지속되고 있으며 실제로 밀과 감자에서는 선별이 성공적으로 이루어진 사례가 있다. 밀의 경우 야생의 Leymus racemosus로부터 유래한 염색체를 도입한 결과 포장에서 고온저항성이 증가한 것을 확인하였다. 벼의 경우에도, 고온과 가뭄 내성 품종인 N22 (aus ssp.)로부터 유래한 QTL이 야간의 고온에 대해 저항성을 부여한다는 것을 밝혀냈다. 이 QTL은 꽃밥 (anther)과 잔이삭 (spikelet), 지엽 (flag leaf)에서의 HSP 고발현과 관련이 있다. 또한 Oryza officinalis로부터 유래한 QTL은 고온에 의한 장해 (spikelet sterility) 회피에 역할을 하는데, 이는 화분 주머니의 열개 (dehiscence)와 수정이 선선한 아침에 일어나게 해주기 때문이다. 이러한 여러 형질의 발견은 가뭄과 함께 고온에 대한 저항성을 증가시킬 수 있는 길을 제시해준다.
9. 맺음말
이상으로 여러 가지 환경에 적응하기 위한 작물들의 다양한 기작 및 이에 관여하는 유전형질을 알아보았다. 환경에 대한 내성을 가지는 여러 유전자들은 진화과정에서 자연변이를 통해 그 특성을 강화시켜 왔다. 이런 유전자원들을 육종 등을 통하여 작물에 도입하는 방법은 각종 규제로부터 자유롭다. 하지만 이렇게 자연적으로 확보할 수 있는 우수 유전형질은 다양성 면에서 제한적일 수 밖에 없다. 그렇기 때문에 위에서 살펴본 여러 유전자들을 유전공학적으로 조절하여 (예컨대, 특정 시간이나 조건에서의 세밀한 발현 조절 등) 작물에 도입하는 것이 대안이 될 수가 있다. 최근 유전자가위로 잘 알려진 tranion activator-like effector nuclease (TALEN)이나 CRISPR-Cas9을 통한 특정 유전부위의 개량 또한 환경스트레스 저항성 부여를 위한 유용한 전략이 될 수 있을 것이다. 이렇듯 분자적 기작에 기반을 둔 작물의 유전공학적 개량이 현재의 식량안보 이슈를 해결할 수 있는 중요한 방법이 될 것이다.
☞ 자세한 내용은 내용바로가기 또는 첨부파일을 이용하시기 바랍니다.
지식
동향