

기술동향
식물 안의 숨겨진 세상: 내생미생물의 기능을 정의하는데 있어서 고려해야 할 생태학적 그리고 진화학적 요인들에 관하여
- 등록일2016-02-03
- 조회수11658
- 분류기술동향
-
자료발간일
2016-02-02
-
출처
생물학연구정보센터(BRIC)
- 원문링크
-
키워드
#내생미생물#생태학#진화학#식물#Microbiology
- 첨부파일
 pdf_0002465.pdf
pdf_0002465.pdf
출처 : 생물학연구정보센터(BRIC)
식물 안의 숨겨진 세상: 내생미생물의 기능을 정의하는데 있어서 고려해야 할 생태학적
그리고 진화학적 요인들에 관하여
?본 자료는 The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev., 2015; 79(3): 293-320 의 논문을 한글로 번역, 요약한 자료입니다.
?
목차
1. 서론
2. 내생미생물 정의의 역사
3. 식물 미생물의 공생
4. 내생미생물의 다양성
5. 내생미생물의 생활사
6. Endosphere의 colonization
7. 내생미생물의 기능
8. 비교유전체 분석을 통한 내생미생물의 특징 분석
9. 병원균과 내생미생물
10. 식물의 생물군계(biome)와 다자간 상호작용
11. 맺음말
1. 서론
내생미생물은 식물체 내에서 전체 생활사의 적어도 일부를 보내는 미생물을 일컫는다. 이러한 정의는 지난 몇 년간 변화되어 왔고 앞으로도 계속 바뀔 것으로 예상이 된다. 처음에 내생미생물(endophyte)이라는 용어는 식물체 내에서 사는 곰팡이들을 지칭하는 용어로 사용이 되었으나, 이후에 곰팡이뿐만 아니라 세균 또한 식물체 내에서 살고 있다는 것이 확인 되었다. 식물의 microbiome이라는 개념이 생기면서 식물의 진화가 단순히 식물의 유전체 자체뿐만 아니라 식물과 연관이 되어 있는 미생물 군집에 대한 선택압(ive force)에 의해서도 영향을 받을 수 있다는 새로운 생각이 나타나기 시작했다 (hologenome concept).
가장 일반적인 내생미생물에 대한 정의는 1997년에 Hallmann과 공동연구자들의 연구로부터 나왔다. 그들은 표면 살균을 한 식물 조직에서 분리되었거나 식물 내부로부터 추출될 수 있는 미생물로서 식물에 눈에 띄는 해를 입히지 않는 것을 내생미생물이라고 정의하고 있다. 그러나 내생미생물의 특성에 대한 개념적인 측면들은 아직 완전히 정립이 되어 있지 않은 상태이다. 예를 들어서, 현재의 개념으로는, 식물병원균들은 그들이 병원성을 잃었을 때에는 내생미생물로 간주를 해야 하는지 아닌지 확실치 않다. 최근에는 한 식물에 대해서 유익한 미생물이라고 할지라도 다른 식물 종으로 갔을 때에는 전혀 상반되는 결과를 주는 경우들이 많이 보고 되고 있다. 또한 가장 흔하게 발견되는 내생미생물들의 경우에 기주식물에 해도 주지 않고 도움도 주지 않는 편리공생의 형태인 경우가 많다. 그러나 이러한 특성들은 일부 종의 식물에 한정되어서만 연구가 되어왔고, 또한 내생미생물 군집 내에서 내생미생물들 사이의 상호작용에 대해서는 거의 연구가 이루어지지 않은 상황이다. 결국 현재 우리의 내생미생물에 대한 이해는 잘 조절되는 조건에서 제한적으로 이루어진 실험결과에 기반한 것이고, 내생미생물의 식물 내에서의 기능과 관계를 보다 깊이 이해하려면 다양한 조건을 전제로 다양한 실험들이 뒷받침 되어야 한다.
내생미생물에 대한 연구는 배양이 가능한 미생물을 위주로 이루어져 온 것 또한 사실이며, 최근에 차세대 염기서열 분석 기술을 통해서 이러한 부분을 보완할 수 있게 되었다. 메타지놈 분석을 통해서 기존에 알려지지 않았던 식물체 내에서 내생세균의 생존에 필요한 기능들을 밝혀낸 연구가 그러한 예가 될 것이다. 이러한 기술적인 발전을 통해서 우리는 내생미생물 연구의 큰 전환점에 서 있다고 하겠다. 앞으로 기술적인 혁신을 통해서 식물체 내의 서로 다른 조직에 존재하는 내생미생물을 식물과의 관계 속에서 보다 잘 이해할 수 있는 기회를 가질 수 있을 것이다.
이 리뷰에서는 내생미생물 연구의 역사적인 측면을 조명하고, 몇 몇의 내생미생물들에 대해서 지난 30여년간 얼마만큼의 연구가 이루어져 왔는지를 살펴보고자 한다. 이와 더불어 앞서 이야기한 기술적인 발전이 앞으로 내생미생물 연구에 어떤 파급효과를 가질 수 있는지에 대해서도 요약을 하고 앞으로 연구의 방향에 대해서 논의하고자 한다.
2. 내생미생물 정의의 역사
내생미생물은 1809년에 처음으로 독일의 식물학자인 Heinrich Friedrich Link에 의해 기술되었다. 그 당시엔 Endophytae라는 용어가 식물에 사는 부분적으로 기생성을 가지고 있는 일부 그룹의 곰팡이를 지칭하는 용어로서 사용이 되었다. 일반적으로 19세기에는 건강하고 정상적으로 자라는 식물은 미생물을 전혀 가지고 있지 않을 것으로 생각을 하였다 (파스퇴르에 의해 제안되었음). 일부 연구자들이 19세기 후반과 20세기 초반에 식물체 내에 식물에 도움을 주는 미생물들의 존재를 확인하였음에도 정상적인 식물은 그 내부에 미생물을 가지지 않으리라는 생각이 그 당시에 계속 유지 되었다.
그 와중에 역사적으로 중요한 발견들이 이루어졌다. 1888년에 네덜란드의 미생물학자인 Martinus Willem Beijerink는 콩과 식물로부터 순수배양을 통해서 뿌리혹 박테리아를 분리하였고, 이어서 이들이 대기 중의 질소를 고정할 수 있는 능력을 가지고 있음을 보여주었다. Albert Bernhard Frank는 또 다른 중요한 상리공생 관계인 나무의 뿌리와 곰팡이 사이의 관계를 보고하였다. 그는 이 관계에 대해서 말 그대로 곰팡이 뿌리를 뜻하는 균근(mycorrhizae)이라는 용어를 처음으로 사용하였다.
좀 더 최근인 1991년에 Orlando Petrini는 내생미생물을 기주식물에 해를 끼치지 않고 식물의 내부 조직들에서 자라며 생활사 중의 일부를 보내는 모든 미생물이라고 정의하였다. 이후에 몇 가지 내생미생물의 정의들이 많은 연구에 기반하고, 실제적으로 내생미생물을 병원균과는 차별화하려는 노력으로 만들어졌으나, 이들은 모두 몇 가지 단점들을 가지고 있고 논란의 여지를 가지고 있다.
우선, 이 정의들은 배양이 가능하고 그래서 병원성 실험이 가능한 내생미생물의 경우에만 적합한 정의들이다. 두 번째로는 일부 세균들은 잠복 병원균 (latent pathogen)으로 식물 내에서 살 수 있으며 다른 식물 종으로 갔을 때에는 이들이 특정한 조건하에서는 병원균이 될 수 있다는 것이 잘 알려져 있다. 세 번째로, 병원균으로서 잘 알려진 미생물이라고 할지라도 다른 식물에서는 생장촉진 효과를 가지는 경우가 있다. 이러한 예들은 내생미생물을 일반적인 병원균과 분리해서 생각한다는 것이 어렵다는 것을 보여주고 동시에 병원성이라든지 공생이라든지 하는 특성들이 정해져 있는 것이 아니라 다른 여러 가지 요인들에 의해서 결정될 수 있는 것이라는 점을 분명히 보여준다. 결론적으로, 현재 사용이 되고 있는 내생미생물의 정의는 굉장히 불완전하며, 따라서 내생미생물이라는 용어는 미생물의 기능이 아니라 서식처(habitat)를 지칭하는 뜻으로만 사용을 해야 할 것이다. 이러한 정의는 식물의 내부 조직들에서 최소한 생활사의 일부를 살 수 있는 모든 미생물들을 포함하게 될 것이다.
3. 식물과 미생물간의 공생관계
뿌리혹 박테리아와 균근은 일부 내생미생물들이 그들의 기주 식물과 공진화했음을 보여주는 예이다. 상호작용에 관여하는 두 파트너의 환경적응력을 높여주는 이러한 상리공생(mutualistic interaction)은 때때로 두 파트너 이상을 포함하는 훨씬 더 복잡한 형태로 진화하기도 한다.
식물-곰팡이 공생의 진화
식물-곰팡이 공생은 육생식물(terrestrial plant)이 지표면을 뒤덮기 시작한 초기에 시작이 되었다고 알려져 있다. Glomeromycota에 속하는 곰팡이가 오르도비스기(Ordovician era)에 첫 육생식물과 상호작용을 한 곰팡이로 생각이 되어왔다. 그러나 Mucoromycotina에 속하는 곰팡이 일부도 첫 육생식물과 공생 관계를 가진 것으로 추측이 된다. 균근과 식물사이의 관계는 첫 육생식물의 육지에서의 적응을 곰팡이가 도와주면서 공생관계로 진화하였다. 그러나 초기에는 식물과 곰팡이 사이에 지금과 같은 밀접한 관계가 형성이 되지는 않았을 것으로 생각된다. 여전히 첫 균근 곰팡이가 이미 상리공생 관계를 가질 수 있는 상태에서 공생을 시작하게 된 것인지 병원성 곰팡이로 시작을 해서 상리공생의 형태로 진화를 한 것인지는 분명하지 않은 상태이다. 어떻든 간에 식물 내부의 공간은 식물을 기반으로 살아가는 곰팡이들에게는 중요한 서식지를 제공해 주었다는 것은 분명하다. 이 과정에서 식물의 외피와 내피 조직이, 뿌리조직으로 내화된(internalized) 곰팡이를 둘러싸는 피질(cortex)의 경계를 형성하도록 진화하면서 균근에서 특징적으로 발견되는 arbuscule이라고 하는 구조가 형성이 되었다. 균근 곰팡이는 시간이 지나면서 좀 더 기주 식물에게 영양과 에너지 면에서 의존적으로 진화하면서 절대활물성 생활사(obligate life cycle)를 가지게 되었다.
식물-세균 공생관계의 진화
가장 연구가 잘 이루어진 식물과 세균 간의 상호작용은 콩과 식물과 근류균(rhizobia)사이의 관계이다. 근류균은 Azorhizobium, Bradyrhizobium, Ensifer, Mesorhizobium, Rhizobium, 그리고 Sinorhizobium 속에 속하는 세균들을 포함하고 있다. 근류균에 대한 전형적인 반응은 뿌리털의 감김(curling)과 감염사(infection thread) 그리고 마지막으로 뿌리의 안쪽 층에서 시원체(primordial)가 나타나는 것이다. 시원체 세포에서는 세균들이 식물의 세포막에 둘러싸이게 되고 식물과 세균이 함께 symbiosome을 형성하게 되는데 바로 이곳에서 대기 중 질소가 고정이 되고 식물의 탄수화물과 맞교환이 이루어지게 된다. 흥미로운 점은 콩과 식물에서 이렇게 뿌리혹을 형성하는 근류균들이 외떡잎 식물들에서는 내생미생물로 발견이 된다는 점이다. 뿌리혹 시원체가 발견이 되지는 않지만 근류균의 nifH 유전자가 발현이 된다는 것은 알려져 있다. 콩과 식물 외의 식물에서 이러한 근류균의 역할과 중요성은 아직 밝혀지지 않았다.
4. 내생미생물의 다양성
원핵(prokaryotic) 내생미생물
현재까지 알려진 모든 16S rRNA 유전자를 저장하고 있는 데이터베이스(저자들이 만든 데이터베이스임)를 기반으로 원핵 내생미생물의 다양성에 대한 조망을 하고자 한다. 신뢰할 수 있는 실험절차를 따른 연구들로부터 나온 300bp이상 되는 염기서열만을 사용하여 분석을 진행하였다. 이 데이터베이스는 순수 분리된 원핵미생물 4,416개에 해당하는 16S rRNA 유전자 염기서열과 3,210 개의 배양이 되지 않는 미생물로부터 얻어진 16S rRNA 유전자를 포함하고 있다. 이들은 23개의 문(phylum)에 속하는 것으로 분석이 되었다 (고세균 2개문, 세균 23개문) (http://mmbr.asm.org/content/79/3/293/T1.expansion.html). 이렇게 놀라울 정도의 다양성에도 불구하고 96% 이상의 염기서열들이 4개의 문에 해당하는 염기서열들이었다(54% Proteobacteria, 20% Actinobacteria, 16% Firmicutes, 그리고 6% Bacteroidetes). 이 문들은 식물에서 우점하는 문으로도 알려져 있다.
대부분의 원핵 내생미생물들이(26%) Gammaproteobacteria에 속하였다. 주목할만한 점은 상당수의 Gammaproteobacteria에 속하는 속(genera)과 종(species)들이 식물병원균으로 알려져 있다는 것이다. 내생미생물로 알려진 Gammaproteobacteria는 주로 Pseudomonas, Enterobacter, Pantoea, Stenotrophomonas, Acinetobacter, 그리고 Serratia 등의 몇 개의 속에서 발견되었다.
전체적으로 보면, 대부분의 내생세균들은 주로 4개의 문에 속하지만, 그 내에서는 다양한 속과 종에 속하는 것을 알 수 있다. 그리고 내생세균들의 기능은 분류에 따라서 달라지는 것이 아니라 기주식물과 환경 요인들에 따라서 달라지는 것으로 보인다.
진핵(eukaryotic) 내생미생물
진핵 내생미생물의 ITS(internal transcribed spacer) 지역에 대한 데이터도 만들어서 분석을 수행하였다. 총 8,439개의 염기서열이 NCBI(National Center for Biotechnology Information)로부터 수집되었다. 이 염기서열들은 주로 취균문(Glomeromycota)(40%), 자낭균문(Ascomycota)(31%), 담자균문(Basidiomycota)(20%), 미확인문(unidentified phyla)(8%), 그리고 접합균문(Zygomycota)(0.1%) 에 해당하였다 (http://mmbr.asm.org/content/79/3/293/T2.expansion.html).
취균문은 균근(mycorrhizae)로 알려진 내생미생물만을 구성하고 있다. 대부분의 진핵 내생미생물은(39%) Glomeromyces 강(class)에 속하였다. 이 강의 모든 멤버들은 대부분의 육상식물들과 내생공생(endosymbiosis) 관계에 있고 따라서 부인할 수 없는 생태적 경제적 중요성을 가지고 있다. 자낭균문 중에서는, 많은 수의 내생미생물이(15%) Dothideomycetes 과에 해당하였다. 내생미생물 외에도 Dothideomycete에 속하는 많은 곰팡이들이 사물기생성(necrotrophic) 식물병원성 곰팡이인데, 이는 이들이 기주 특이적인 독소를 생성할 수 있다는 점에서 주목할 만한 점이다.
Sordariomycete과에 속하는 많은 곰팡이들도(9%) 내생곰팡이였다. Balansia, Epichloe, Nemania, Xylaria 그리고 Colletotrichum 속에 속하는 종들이 이에 해당한다. 이 과에 속하는 곰팡이들 역시 상당수가 Cryphonectria parasitica, Magnaporthe grisea, Ophiostoma ulmi, 그리고 Fusarium, Verticillium, Rosellinia종에 속하는 식물병원균으로 알려져 있다. 담자균문 중에서는, Agaricomycete과에 속하는 곰팡이가 많은 수의 내생곰팡이(18%)를 포함하고 있었는데, 이들은 버섯을 형성하는 곰팡이, white-brown rot 부생균 등에 해당한다.
내생세균에서와 마찬가지로 내생곰팡이가 속하는 다양한 분류군들이 식물병원균뿐만 아니라 병원성이 알려지지 않은 균주들을 포함하고 있었다. 이는 다시 한 번 내생곰팡이의 기능이 반드시 분류상의 위치와 상관관계가 있는 것은 아니라는 것을 보여준다.
5. 내생미생물의 생활사
생활사를 완성하기 위해서 반드시 식물의 조직을 필요로 하는 미생물을 절대활물성 (obli-gate)이라고 한다. 연구가 잘 되어 있는 절대활물성 내생미생물에는 균근(mycorrhizae)과 Clavicipi-taceae과에 속하는 Balansia, Epichloe, 그리고 Neotyphodium 속(genera)의 멤버들이 해당한다. 절대활물성의 다른 끝에는 식물 조직 밖에서(epiphyte) 주로 번식하며 산발적으로 식물의 en-dosphere로 들어오는 기회성(opportunistic) 내생미생물들이 있다. 이들 중에는 Pseudomonas, Azospirillum 속(genera)에 속하는 세균들과 Hypocrea와 Trichoderma속(genera)에 속하는 곰팡이들이 있다.
이 양 극단의 중간에, 대부분의 내생미생물들을 포함하고 있는, 조건적(facultative) 내생미생물군이 있다. 이들 조건적 내생미생물들이 식물을 전파(dissemination)를 위한 매개체로 쓰고 있는 것인지 아니면 이들 미생물들이 식물에 의해서 적극적으로 선택되는 것인지는 분명치 않다. 그러나 조건적 내생미생물들은 식물이 주는 영양분들을 소비하고 있고, 이는 기주식물의 적합성(fitness)를 사실상 감소시키게 된다.
이들 세 개의 그룹 사이에는 겹치는 부분들이 존재하기 때문에 이러한 구분은 내생미생물들 사이에 존재하는 식물 내 colonization전략의 연장선 상에 임의적으로 경계를 지은 것으로 생각을 해야한다.
6. Endosphere의 colonization
내생곰팡이의 colonization 특성
내생미생물의 식물 내에서 성공적인 생장은 많은 변수들에 의해서 결정이 된다. 이러한 변수들에는 식물 조직의 종류, 식물의 유전형, 미생물의 종류, 그 외의 생물학적인 조건과 환경 조건들이 있다. Clavicipitaceae와 nonclavicipitaceae 내생미생물의 다양한 식물 내colonization 전략이 알려져 있다. Clavicipitaceae에 속하는 Balansia spp., Epichloe spp., 그리고 Claviceps spp.들은 거의 예외없이 잔디와 같은 기주와 공생 관계를 맺고 있으며, 특정 기관이나 조직이 아니라 식물 전체에서 colonization을 하는 특성을 가지고 있다. 이들은 식물의 씨앗을 통해서도 수직전반(vertical transmission)을 할 수도 있다. 일부 Neotyphodium이나 Epichloe 종들은 땅에 떨어지는 잎 조각들을 통해서 수평전반(horizontal transmission)을 하기도 한다.
내생미생물들의 생장 특성에 따라서 Rodriguez 등은 clavicipitaceous에 속하는 내생미생물들을 class 1 내생 곰팡이로 분류하였다. 식물의 지상부 및 지하부 즉 근권을 coloniza-tion을 하고 수직 그리고/또는 수평 전반을 하는 곰팡이들은 class 2로 분류하였다. Class 3에 속하는 내생곰팡이들은 대부분이 Dikaryomycota에 속하는 곰팡이들로 이들은 수목과 관련해서 연구가 특히 잘 되어 있으나 다른 식물 분류군에서도 연구가 되어 있기는 하다. 이 class에 속하는 곰팡이들은 대체로 식물의 지상부에 그 범위가 한정이 되어 있고, 주로 수평전반을 한다. Class 4에 속하는 내생곰팡이들은 어두운, 격벽이 있는 내생곰팡이들로서, 근균(mycorrhizae)과 유사하게 뿌리 조직으로 그 범위가 제한되어 있다.
내생세균의 colonization특성
많은 내생세균들은 식물이 뿌리 삼출물(exudate)로 미생물을 유인하게 되는 근권 환경으로부터 유래한다. Mercado-Blanco와 Prieto는 내생세균들이 식물 뿌리털의 colonization을 통해서 식물 내부로 들어올 것이라고 주장하였다. 식물의 줄기나 잎 부분도 어느 정도는 삼출물을 생산하기는 하지만, 자외선을 받게 되고, 영양소와 수분이 부족한 환경은 일반적으로 미생물이 식물 잎 위를 colonization하는 것을 저해하게 된다.
균주의 종류에 따라서 다양한 colonization 루트가 있다는 것도 보고가 되었다. 이러한 루트에는 수동적 그리고 능동적 메커니즘이 있으며 이들은 모두 세균이 근면(rhizoplane)에서 뿌리의 피층(cortical layer)으로의 이동을 가능하게 한다. 내피를 관통해서 들어갈 수 있는 세균의 경우에는 물관 시스템을 통해서 전신성(systemic) colonization을 하게 되고, 반면에 다른 세균들의 경우에는 식물세포 사이의 공간들을 국지적으로 colonization하게 된다. 내생세균들은 특정한 기관이나 조직에서 발견이 되기도 하는데, 그러한 조직으로 이동하는 것이 내생세균의 식물 내에서의 기능을 위해서 필요한 것인지 아닌지에 대해서는 더 많은 연구가 필요하다.
7. 내생미생물의 기능
일부 내생미생물들은 식물에는 거의 영향을 주지 않고 식물이 만드는 대사 산물들을 기반으로 살아가기도 한다. 반면에 다른 내생미생물들은 식물을 병원균이나 초식동물(arthropod herbivore)로부터 보호해 주거나 식물의 생장을 촉진해 줌으로서 도움을 주기도 한다. 이 외에도 잠재적 병원균 그룹이 있다. 일반적으로 내생미생물이 식물에 미치는 영향은 여러 가지 다른 조건들의 조합에 의해서 결정이 된다. 예를 들어, Fusarium verticillioides는 병원균과 유익한 내생미생물의 두 가지 기능을 모두 가지고 있다. 이 두 상태 사이의 균형은 기주 식물의 유전형 및 기주 식물의 적합성(fitness)에 영향을 줄 수 있는 비생물학적인 환경 조건에 의해서 결정이 된다. 현재까지 알려진 내생미생물의 다양한 기능들은 아래와 같이 요약할 수 있다 (그림 1).
식물생장촉진과 생물학적 비생물학적 스트레스로부터의 보호기능
Carroll 등은 1991년에 내생미생물에 의해서 식물에 전신유도저항성(induced systemic re-sistance)가 생길 수 있다는 것을 보여주었다. 따라서 내생미생물을 가지고 있는 식물은 그렇지 않은 식물에 비해서 더 높은 수준의 병원균에 대한 저항성 및 내성을 가질 수 있다. 이러한 내생미생물에 의한 식물의 면역반응 유도에 대한 증거는 점점 많아지고 있다. 이러한 유도된 면역반응은 병원균이 침입했을 때와 유사하나 상리공생을 하는 내생미생물들은 이러한 면역반응을 피하여 성공적으로 기주식물을 colonization할 수 있다는 것도 밝혀졌다. 내생세균의 경우에는 세균의 편모(flagella), N-acetylhomoserine lactone, salicylic acids, jasmonic acids, siderophore와 같은 요인들이 전신유도저항성을 일으킬 수 있는 것으로 밝혀졌다. 흥미로운 점은 이렇게 저항성이 유도가 될 때에는 식물 내에 있는 내생미생물의 군집 구조에도 변화가 생긴다는 점이다. 이러한 군집 구조의 변화는 병 저항성과 상관관계가 있음이 밝혀졌는데, 이는 내생미생물의 군집이, 최소한 그 일부라고 할지라도, 병 발생의 억제에 관련성이 있음을 시사한다. 내생세균과는 다르게 내생곰팡이의 경우에는 전신유도저항성에 관련이 있다는 보고는 훨씬 더 적었다.
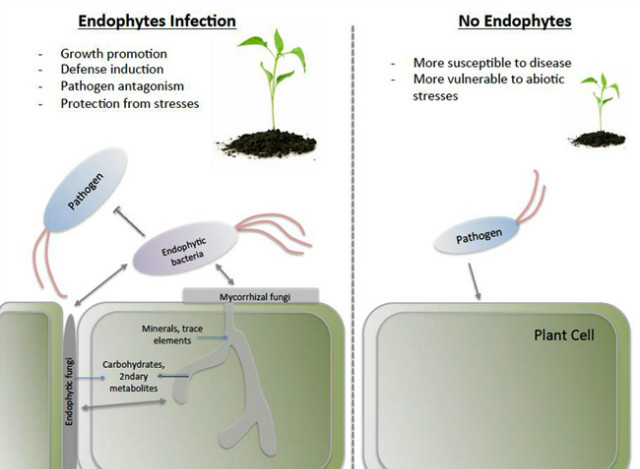
그림 1. 내생미생물의 기능들.
내생미생물이 존재할 때와(왼쪽) 존재하지 않을 때(오른쪽)의 상황을 모식도로 비교하였다. 회색 화살표 및 선들은 미생물간의 상호작용을 파란색 화살표는 물질의 이동을 표시한다.
내생곰팡이들은 식물병원균이나 초식동물에 대해서 생장억제 효과를 가지는 물질을 생성하는 능력으로 더 잘 알려져 있다. 이러한 물질들에는 알칼로이드, 스테로이드, 터피노이드, 펩타이드, 폴리키톤, 플라보노이드, 퀴놀, 페놀계 물질들이 있다. 알칼로이드는 clavicipitaceous 내생곰팡이에 의해서 생성되는 물질로서 가장 연구가 많이 되어 있다. 예를 들어, 롤리트렘(lolitrem)이라고 불리는 신경독성이 있는 indole-diterpenoid alkaloid는 내생미생물에 감염되어 있는 풀을 먹는 가축의 중독현상에 관련이 되어있는 물질이다. 최근에는 내생미생물에 의한 항생물질(antimicrobial)의 생성이 식물을 침입하는 병원균에 의해서 유도가 될 수 있다는 것이 알려지고 있다.
내생세균들도 마찬가지로 항생물질들을 생성할 수 있다. 일반적으로, 내생성 방선균(actinomycete)들이 가장 잘 알려진 항생물질의 생산자로서, 이들이 만드는 대표적인 항생물질은 munumbicin, kakadumycin, coronamycin등이 있다. 이 외에도 다양한 물질들이 보고가 되고 있고, 이들이 활성이 의학이나 농업에서 활용가치가 있을 것으로 예상이 되나, 식물과 내생미생물간의 상호 작용에서의 정확한 역할은 아직 알려지지 않은 상태이다.
미생물의 2차 대사산물들은 보통 생물학적 활성이 강해서 항암, 항산화, 항당뇨, 면역억제, 항미생물제, 항바이러스제로서 쓰일 수 있는 가능성을 가지고 있다. 내생미생물들은 신호전달, 방어반응, 공생관계에 관여하는 다양한 2차 대사산물들을 생성하는 것으로 알려져 있다. 단순히 자신이 2차 대사산물들을 생산하는 것을 넘어서 내생미생물들은 기주 식물의 2차 대사산물 생산에도 영향을 줄 수 있다. 예를 들어 딸기를 Methylobacterium 으로 감염시키면 furanone과 같이 딸기의 향에 영향을 주는 물질의 생합성에 영향을 주는 것이 알려져 있다.
일부 내생세균과 내생곰팡이들은 siderophore를 많이 만들기도 한다. Siderophore는 토양 미생물들이 철분(iron)을 얻는데 중요한 역할을 하지만, 동물에서는 기주와 병원균 사이의 상호작용에서도 중요한 역할을 하는 물질이다. 내생미생물이 식물을 colonization 하는데 있어서 siderophore의 역할은 명확하지 않으나 이 물질들이 유도전신저항성을 일으키는데 관여하고 있을 가능성이 있다. 뿐만 아니라 Epichloe festucae의 경우에는 독보리(ryegrass)와의 공생관계에 있어서 중요한 역할을 한다는 것이 밝혀져 있다. 내생성 Methylobacterium가 만드는 siderophore는 감귤(citrus) 나무에서 백화병(chlorosis)을 일으키는 Xylella fastidiosa의 생장을 억제한다는 것도 알려져 있다.
대부분의 내생미생물들이 식물을 생물학적 스트레스로부터 보호하는 기능을 가지는 것이 잘 알려져 있지만, 일부 내생미생물들은 식물을 비생물학적인 스트레스로부터 보호하는 기능도 가지는 것으로 밝혀지고 있다. 예를 들어서, Neotyphodium spp.은 삼투 및 기공 조절을 통해서 식물의 가뭄 내성을 증가시키고, 또한 질소부족이나 물 부족으로부터 식물을 보호하는 기능을 하는 것으로 알려져 있다. 뿌리의 내생곰팡이인 Piriformospora indica는 보리가 염분내성을 가지도록 유도하고 배추에서는 가뭄 내성을 가지도록 해주는 기능을 가지고 있었다. 두 가지 경우 모두에서 항산화제 수준을 증가시키는 것이 이 식물들에서 스트레스에 대한 내성을 올려주는 메커니즘인 것으로 보인다. 이 외에도 Trichoderma나 Burkholderia에 의한 식물의 비생물학적인 스트레스에 대한 내성 증가가 보고된 바 있다.
일부 내생미생물들은 식물의 생장촉진에 관여하기도 한다. 내생미생물들이 식물 내에서 살기 위해서는 식물로부터 영양분을 받아야 한다는 것을 생각할 때에, 식물에서 많은 수의 내생미생물들이 발견된다는 것은 적어도 둘 사이의 관계에서 손익(cost-benefit) 균형이 중립적이거나 식물에 이익이 되는 쪽으로 기울어 있다는 것을 의미한다. 따라서 대부분의 내생미생물들은 유익균일 가능성이 높으며 그러한 유익한 효과(beneficial effect)는 식물의 광합성이나 탄소고정 과정에 내생미생물들이 관여하기 때문인 것으로 생각이 된다. Neotyphodium lolli는 기주식물의 광화학과정이나 광합성 자체에는 영향을 주지 않지만, 이산화탄소 고정률에 영향을 주는 것으로 알려져 있다.
내생미생물에 의해서 생산되는 식물호르몬이 아마도 내생미생물이 식물생장을 촉진할 수 있는 가장 잘 알려진 메커니즘일 것이다. 옥신이나 지베렐린을 생성하는 능력은 식물의 뿌리와 연관이 되어 있는 내생미생물의 전형적인 특징이라고 할 수 있다. Indole-3-acetic acid(IAA)는 기주식물의 방어반응을 조절함으로써 내생미생물의 colonization 효율을 높여주는 역할을 하는 것으로 보인다. 일반적으로 이러한 호르몬들의 생성은 내생미생물이 식물을 colonization하는데 있어서 중요한 역할을 하는 것으로 추측이 된다. 사이토키닌(cytokinin)의 생성도 내생미생물에서 흔하게 발견이 되며, 이러한 사이토키닌 생성능력은 식물생장 촉진과도 밀접하게 관련이 되어 있음이 보고 되었다.
식물호르몬 생산 외에도 내생미생물에 의해서 만들어지는 2,3-butanediol과 같은 휘발성 물질이나 폴리아민류의 물질도 역시 식물의 발달이나 생육에 영향을 주는 것으로 알려져 있다. 또한 질소고정을 통해서 식물의 생육을 촉진하는 것도, 잘 알려져 있는, 내생미생물이 식물생육을 촉진하는 메커니즘이다. 일부 뿌리 내생미생물들이 질소고정을 한다는 것이 알려져 있으나 이들의 효율은 콩과 식물의 뿌리혹에서 일어나는 질소고정에 비하면 효율이 굉장히 낮은 편이다. 이러한 내생미생물의 질소고정 능력은 특히나 질소가 많지 않은 토양 환경에서는 더욱 중요하다고 할 수 있다.
식물의 환경적합성을 향상시키는 미생물과의 공생관계
균근(AMF: arbuscular mycorrhizal fungi)이나 근류균(rhizobia)와는 분류학적으로 다른 내생미생물들도 기주 식물의 환경 적합성(fitness)을 증가시키는 것이 잘 알려져 있다. 예를 들어 반점 뻐국채류(spotted knapweed)를 내생곰팡이인 Alternaria alternata로 접종을 하면 다발풀(bunch grass)과의 관계에서 보다 경쟁력을 가지게 되는 것이 보고되었다. 또 다른 경우에는 왕김의털(red fescue)이라는 식물이 병원균의 하나인 Claviceps purpurea에 감염이 되면 양과 같은 동물들에 의한 섭식이 감소하는 것으로 알려져 있다.
8. 비교유전체 분석을 통한 내생미생물의 특징 분석
비교 유전체학은 내생미생물의 식물 침입과 colonization에 있어서 중요한 유전자들을 발굴하고 연구하는데 있어서 중요한 도구라고 할 수 있다. 분류학적으로 가깝지만 내생성을 가지고 있지 않는 다른 미생물들과의 비교를 통해서 내생미생물이 되기 위해서 필요한 특징들이 무엇인지를 파악할 수 있을 것이다. 최근에 이루어진 비교유전체 및 대사체 연구에서 상리공생을 하는 미생물과 병원균들 사이에 여러 가지 차이들이 있음을 발견되었다. 일반적으로 물질 생합성에 관여하는 유전자들은 상리공생을 하는 미생물들에서 더 많이 더 다양하게 존재하는 것을 알 수 있었고, 병원균에서는 물질 분해 및 기주 식물의 침입과 관련된 유전자들이 상대적으로 더 많음을 알 수 있었다. 세균의 경우에는 유전체의 크기와 기주 범위가 상관관계가 있음도 알 수 있었다.
내생성 세균에서 진화된 특징들을 밝히기 위한 비교유전체 연구
내생미생물의 기능적(functional)인 측면에 대해 좀 더 밝히기 위해 여기에서는 식물 조직 내에서 분리된 40 여 strain의 잘 연구된 세균들을 대상으로 42개의 뿌리혹 박테리아, 29개의 병원성 세균, 그리고 42개의 근권 세균과의 비교 연구를 수행하였다. 단백질을 암호화하는 유전자들에 대해서 Integrated Microbial Genome (IMG) 비교분석 시스템 (http://img.jgi.doe.gov)을 사용하여 KEGG Ortholog(KO) 태그들을 할당을 하고 분석을 진행하였다 (http://mmbr.asm.org/content/79/3/293/T3.expansion.html). 이 분석을 통해서 내생미생물이 가지는 전형적인 특징이라고 할만한 지표들을 찾아내고 이들에 대해 앞으로 연구해 가야 할 방향성을 제시하고자 하였다.
세균의 주화성과 운동성에 관련되어 있는 유전자들에 대한 분석을 통해서 aspar-tate/maltose(Tar) 및 dipeptide(Tap)의 이용에 관련한 유전자들이 근권 세균에서보다 내생세균에 더 많이 존재한다는 것을 알 수 있었다. 반응조절자(response regulator)인 CheBR과 CheC 그리고 편모의 생합성 유전자, 운동성 유전자들도 병원균보다는 내생세균에 더 많이 존재하였다.
Two-component response system 측면에서 내생세균들은 주로 뿌리혹 박테리아들과 많은 차이를 보여주었고, 다른 그룹들과는 큰 차이가 없었다. 항생제 저항성이나 산화환원반응, 질소고정 및 대사, 그리고 세포운명(cell fate)에 관련된 유전자들은 병원균이나 근권 세균에 비해서 내생세균들에서 더 많이 발견되었다.
전사조절인자들에 대해서 분석을 했을 때에는, 질소 동화(nifA), 산화질소의 환원(norR), 탄소 저장 조절(sdiA), 베타락타메이즈 저항성(ampR), 피리미딘 대사(pyrR), 그리고 티아민(thiamine) 대사에 관련된 유전자들이 다른 그룹들보다 내생세균들에서 더 높은 비율로 나오는 것을 알 수 있었다.
식물의 방어 반응으로 만들어지는 활성산소(reactive oxygen species)를 잘 제어하고 해독(detoxification)하는 것도 내생미생물의 성공적인 colonization을 위해서 중요한데, 실제로 glu-tathione peroxidase(btuE)나 glutathione-S-transferase(gst), catalase(katE) 등의 유전자들은 내생세균의 유전체에 풍부하게 존재하는 것을 알 수 있었다. 또한 내생세균들은 다당류, 유기산, 펩타이드, 아미노산, 탄수화물의 흡수에 관련된 유전자들을 다른 그룹의 세균들보다 더 많이 가지고 있었다.
이뿐만 아니라 단백질의 분비에 관련된 유전자들은 내생세균이 근권 세균들보다는 많이 가지고 있는 경향을 보였다. 그러나 그 숫자가 뿌리혹 박테리아나 병원성 세균들에서 관찰되는 숫자보다는 적었다.
질소 고정에 관여하는 유전자들은 병원성 세균보다 확실히 내생세균들에서 훨씬 더 많은 숫자가 존재하고 있었으며, 놀랍게도 조사된 모든 내생세균의 약 28% 정도가 이 유전자(nifH)를 가지고 있었다. 이는 질소가 제한되는 환경조건에서 식물의 생산성을 올리는데 있어서 내생세균들이 중요한 역할을 한다는 것을 보여주는 것이라 하겠다.
요약하자면 이러한 비교유전체 분석을 통해서 여러 기능적으로 다른 그룹의 세균들 사이에 차이가 존재한다는 것을 알 수 있었다. 그러나 이러한 경계는 분명히 나누어지는 것이 아니며 기능적인 그룹들 사이에 많은 겹치는 부분들이 있다는 것을 인식해야 한다. 그럼에도 불구하고 내생세균들은 분명히 다른 그룹에 비해서 환경변화, 질소고정, 그리고 활성산소 등에 대해서 더 민감하게 반응할 수 있는 특징들을 가지고 있다는 것은 분명한 사실이다.
9. 병원균과 내생미생물: 상호작용에서 균형의 중요성
병원성 ? 정의와 메커니즘
병원성은 미생물이 가지는 가장 눈에 띄는 특성 중의 하나이다. 최근에 차세대 유전체 염기서열 분석 기술을 통해서 인간의 미생물군유전체(microbiome)와 미생물의 병원성에 대한 우리의 지식이 폭발적으로 증가하고 있다. 특히 사람의 미생물군유전체에 대한 연구로부터 미생물군유전체가 기존에 생각했던 것보다 훨씬 더 많은 질병의 발생에 관여하고 있다는 것과 병원균의 인체 내에서의 갑작스런 증식과 뒤따르는 질병이 전체 미생물 군집의 변화와 연관성을 가진다는 것을 알게 되었다.
일반적으로 사용되는 내생미생물의 정의는 병원성 미생물 자체를 포함하지는 않는다. 그러나 모든 최근의 연구들은 식물과 내생미생물의 상호작용이 기주 식물에 유익한 것에서부터 병원성에 이르기까지 훨씬 더 넓은 범위에 걸쳐 있다는 것을 보여주고 있다. 많은 인체병원균들뿐만 아니라 식물병원균들이 식물의 endosphere를 colonization 할 수 있다는 것이 알려져 있다. 따라서 내생미생물들과 내부병원균(internal pathogen)들은 몇 가지 메커니즘을 공통적으로 가지고 있다. 그러한 예에는 siderophore라든지 제3형 분비시스템(Type III secretion system)이 있다.
식물의 endosphere 내의 잠재적 병원균의 발생(occurrence)
식물의 endosphere는 식물, 동물, 그리고 인간의 병원균들에 의해서 colonization이 될 수 있다. 병원성에 의해서 병징이 나타나지는 않았으나, 실제로 식물의 미생물군유전체에 대한 분석에서 그러한 병원균들이 분리되었다. 내생곰팡이인 Verticillium dahliae는 한 가지 흥미로운 예이다. 원래 이 곰팡이는 여러 종류의 작물에서 수확량 손실을 초래하는 병원균이다. 그러나 한편으로 이 곰팡이는 많은 건전한 식물에서 편리공생하는 내생미생물로서 발견이 된다. 동물병원균이 병원성 대장균도 endosphere를 colonization 하는 것을 확인할 수 있었다. 기회성 병원균들은 특히 주목할만한데 그 이유는 이들이 식물의 endosphere 내에 존재하면서 식물이 이들 병원균들의 저장고(reservoir)로서 기능할 수 있기 때문이다.
식물의 잎을 가해하는 병원균에 대한 저항성은 일반적으로는 식물의 유전체상에 암호화되어 있으나 식물과 밀접한 관계를 가지는 미생물들에 의해서 매개되어지기도 한다. 특히 뿌리 병원균에 대한 저항성 유전자는 상대적으로 드문데, Cook 등은 이러한 기능을 근권세균이 대신해줄 수 있을 것이라고 이야기하고 있다.
10. 식물의 생물군계(biome)와 다자간 상호작용
내생미생물의 군집 구조
식물의 뿌리는 식물이 환경 및 미생물과 상호작용을 하고 신호등을 교환하는 장소라는 점에서 흔히 사람의 장(gut)에 비유가 된다. 이러한 생각은 인간의 장내 미생물 군집의 구조와 식물의 뿌리의 미생물 군집 구조를 비교하는 연구로 이어지게 되었다. 식물의 내생미생물의 군집구성은 생물학적 비생물학적 요인들 모두에 의해서 조절이 된다. 가장 중요한 것은 식물 즉 기주의 유전형과 발달단계 그리고 내생미생물이 유래하는 환경 조건이다. 최근 분자수준 군집분석(molecular community analysis) 결과를 보면 토양의 타입이 식물의 유전형보다도 세균 군집의 조성을 결정하는 가장 중요한 요인인 것을 알 수 있다. 일반적으로 유전적으로 가까운 식물들은 유사한 내생미생물 군집 조성을 가지는 것이 보고가 되고 있으나, 기주 식물들 사이의 계통분류학적인 거리가 세균의 군집 구성의 다양화를 완전히 설명해 주지는 못한다.
내생곰팡이는 종자전반(seed transmission)을 하는 것이 잘 알려져 있다. 최근에는 내생세균들도 종자전반을 할 수 있는 가능성이 제기되었다. 종자에서 발견되는 내생세균 군집은 토양환경에서 발견되는 군집과는 다소 독립적이고 다른 것으로 보고 되었다. 이는 수직전반(vertical transmission)되는 내생세균들 또한 식물의 환경적합성(fitness)에 기여할 수 있는 가능성을 제시하는 것이다.
내생미생물의 다자간(multitrophic) 상호작용
식물의 생물군계(biome)는 식물과 다양한 곰팡이 및 세균들로 구성되어 있다. 이들은 아주 복잡하고 조밀한 네트워크를 구성하고 있다. 그러나 이러한 구성원들 사이의 상호작용과 그로부터 만들어지는 네트워크는 거의 연구가 되어 있지 않은 상태이다. 특히 식물과 내생미생물들 또는 내생미생물들 사이의 밀접한 관계, 신호전달, 인식 과정은 연구가 부족한 상태이다. 식물과 내생미생물간의 상호작용은 식물과 다른 미생물간의 상호작용에도 영향을 준다는 점에서 이 과정에 대한 이해는 중요하다고 하겠다. 내생미생물이 식물 병원균에 대해서 길항작용을 할 수 있다는 것은 알려져 있으나 구체적인 메커니즘은 확실히 밝혀져 있지 않다. 내생미생물에 의한 항생효과는 복잡한 화학적 커뮤니케이션 (chemical communication)에 의해서 유도가 되는 것일 수도 있다. 예를 들어서 항생효과가 있는 내생미생물을 독립적으로 배양한 후에 그 추출액을 처리하면 아무런 효과가 없지만 내생미생물을 병원균과 함께 배양하면 항생효과나 다른 효과가 있는 대사물질들의 생산을 유도할 수도 있는 것이다. Paraconiothyrium variable은 식물의 잎 조직에서 발견되는 내생곰팡이로 이들은 Fusarium이 존재할 때에 옥시리핀(oxylipin)이라는 물질을 생산을 하고, 이는 Fusarium의 진균독소(mycotoxin)의 생성을 억제하는 효과를 갖는다. 이러한 예는 식물과 미생물뿐만 아니라 미생물과 미생물 사이의 화학적 커뮤니케이션이 중요함을 잘 보여주고 있다. 그러나 우리는 아직 이제야 다자간 상호작용을 연구하고 이해하기 시작하고 있다.
내생미생물은 곤충을 포함하는 동물들에 대한 식물의 방어를 증가시키는 역할을 할 수도 있음이 알려져 있다. 그러한 한가지 메커니즘은 내생미생물에 의한 알칼로이드 생성인데, 이는 식물병원성 바이러스의 매개충인 진딧물의 식물에 대한 공격을 감소시키는 것이 보고 되었다. 내생미생물의 군집 구조는 거꾸로 병원균에 의한 식물의 감염에 의해서도 영향을 받을 수 있다. 이는 잠재적으로는 미생물의 기능에 영향을 주게 된다. 일부 연구들은 내생미생물이나 내생곰팡이의 다양성 감소가 이미 병들었거나 병원균을 내재하고 있는 식물들에서 일어나고 있음을 보고하였다. 이는 아마도 병원균에 의한 감염이 식물에서 스트레스와 관련이 있는 대사 산물의 생산을 증가시키고 이것이 기존과는 다른 종류의 생리학적인 특성을 갖는 서식처를 만들기 때문인 것으로 추정해 볼 수 있다. 내생미생물과 병원균과의 상호작용과 별도로, 내생미생물들은 다른 공생미생물들과도 상호작용을 한다. 잎에서 발견되는 내생곰팡이들의 조성은 균근곰팡이의 colonization에 의해서 변화가 된다는 것이 보고 되었다. 결론적으로, 식물의 생물군계(biome)는 식물과 다양한 기능의 미생물 그룹들을 포함하는 다자간의 복잡한 상호작용에 의해서 특징지워짐을 알 수 있다. 식물의 형질은 식물의 환경에 대한 반응에 의해서만 결정되는 것이 아니라 식물과 밀접한 관련성을 가지는 미생물 군집에 의해서도 조절이 된다는 된다는 점을 인식해야 한다.
11. 맺음말
기술적인 발전들 특히 오믹스(-omics) 기술들은 endosphere 내에 존재하는 미생물군유전체에 대한 우리의 기존의 생각과 개념들을 완전히 바꾸어놓게 될 것이다. 이러한 기술들을 통해서 우리는 여전히 알려지지 않은 내생미생물들의 역할을 이해할 필요가 있다. 특히 내생미생물들의 다수를 차지하는 편리공생하는 것으로 알려진 내생미생물들의 역할에 대한 연구가 필요하다. 이들에 대한 연구를 통해서 그 동안 알려지지 않았던 새로운 내생미생물들의 역할을 밝혀낼 수도 있을 것이다. 또한 어떤 조건에서 그리고 어떤 메커니즘으로 내생미생물들이 식물에 해를 끼치게 되는지, 이익을 주게 되는지 아니면 중립적인 효과를 가지게 되는 것인지에 대해서 좀 더 잘 이해할 필요가 있을 것이다. 앞으로 새로운 기술적 발전들과 다학제적인 접근을 통해서 내생미생물들에 대한 우리의 이해를 증진시키고 이를 바탕으로 식물의 전생물계(holobiome)에 대해 좀 더 잘 이해해야 할 것이다.
☞ 자세한 내용은 내용바로가기 또는 첨부파일을 이용하시기 바랍니다.