

기술동향
SMC proteins 2017: Chromosomal organizers from bacteria to human 참관 후기
- 등록일2017-09-19
- 조회수5131
- 분류기술동향 > 종합 > 종합
-
자료발간일
2017-08-31
-
출처
생물학연구정보센터(BRIC)
- 원문링크
-
키워드
#SMC proteins# condensin#염색체 응집#염색체 분열#cohesin
- 첨부파일
 pdf_0002807.pdf
pdf_0002807.pdf
출처 : 생물학연구정보센터(BRIC)
SMC proteins 2017: Chromosomal organizers from bacteria to human
참관 후기
류제경 / 네덜란드 델프트 공과대학교
[목 차]
Ⅰ. 학회 소개
Ⅱ. 주요 발표 내용
1. Keynote session
2. Prokaryotic SMC session
3. Chromosome biophysics and modeling
4. Sister Chromatid cohesion
5. Condensin
6. Chromosome organization in meiosis/zygotes
7. SMC and Disease
Ⅲ. 총평
[요약문]
2017년 6월 13일부터 16일까지 일본 Yamagata 현의 Nanyo시의 Nanyo City Cultural Hall에서 SMC proteins 학회가 Chromosomal organizers from bacteria to human이라는 주제로 진행이 되었다. Oral presentation은 42개 poster는 71개 정도로 이뤄졌고, 인원은 300명 정도 참여한 것 같다. 모든 연구자가 톡을 같이 듣고, 함께 디스커션하고, 함께 먹고 마시면서 Excursion을 즐겼다. 이 학회는 SMC 단백질을 초점으로 열렸고, 아마 생물학 학회 중에서 한 단백질군만 집중적으로 보고하는 학회는 아마 이것이 유일할 것이다. Cell biology, biochemistry, biophysics 분야가 함께 어울러져서 함께 이 단백질에 대한 집중 연구 보고가 이뤄졌다. SMC 단백질의 구조가 어떻게 생겼는지, 어떤 구조 변화를 거치는지, 이 단백질이 Cell 안에서 어떻게 작용하여 염색체를 조작하며 염색체 분열을 일으키는지에 대한 연구들이 함께 연결되어서 좋은 하모니를 이뤘다고 생각한다. 거기에 더 나아가서 어떤 질병을 우리가 해결할 수 있는지도 함께 나눈 학회였다고 생각한다. 함께 식사와 다과를 하면서 이 분야 사람들이 함께 어울러져서 생물학의 과제 중에서 가장 큰 숙제를 함께 풀고 있다는 느낌을 많이 받았다.
Ⅰ. 학회 소개
SMC proteins 학회는 SMC 단백질 family만을 초점으로 둔 학회이다. 첫 SMC protein 학회는 EMBO 학회로 2015년에 유럽에서 열렸고, 이번에는 두 번째로 SMC 단백질 강국인 일본에서 열리게 되었다. Nanyo시는 아주 조그마한 시골마을이었다. 그래서 많은 일본인들도 모르는 지역이었다. 하지만, 아주 평온한 농촌 마을 속에서 평화롭게 학회는 진행되었다.

멀리 보이는 회색 빛 건물이 이번 SMC 단백질 학회가 열렸던 곳이다.
평화로운 시골 농촌 마을에서 열렸고, 학회 내내 날씨는 맑았다.
SMC 단백질은 condensin, cohesin과 SMC5/6로 되어 있으며 chromosome organization과 segregation에 관여하는 단백질이다. 한동안 정제 기술의 어려움으로 cell biology에서 많이 진행되었다가 한동안 뜸했지만, 최근 정제 기술의 발전으로 많은 연구가 진행되고 있다. SMC 단백질의 공통적인 특징은 두 개의 SMC 단백질이 hinge 도메인에 의해서 결합되어 dimer를 만들고, 또 각각 ATPase head domain이 있고, 이 head domain과 hinge는 coiled coil 구조로 연결이 되어 있다. Kleisin subunit은 ATPase head domain 각각에 결합되어 SMC-kleisin ring 구조를 만들어낸다. 그리고 단백질에 따라 여기에 한 두 개의 non-SMC 서브 유닛들이 붙어 있다.
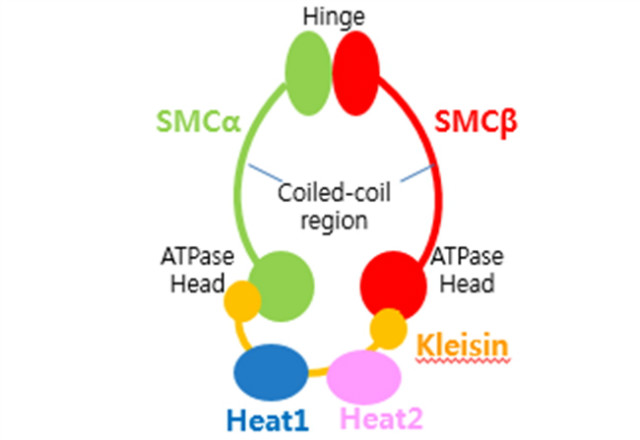
그림 1. SMC 단백질의 공통 구조.
우리가 중학교 때 배우는 세포 분열과 염색체 분열에 관여 하는 key 단백질이지만, 아직도 이 단백질의 기작은 미스터리로 되어 있다. SMC proteins 학회처럼 단백질 군 하나만 집중적으로 다루는 학회도 드물 것이다. 이번 학회는 두 번째 개최되는 학회로 첫 번째 학회는 EMBO meeting으로 진행되었다가 성황리로 마무리 되었고, 이번 학회는 SMC 단백질 연구의 강국인 일본에서 진행되게 되었다. 일본은 Tatsuya Hirano, Katsuhiko Shirahige, Yuri Nakagawa, Mitsuhiro Yanagida 등의 리딩 그룹이 있고, 유럽과 미국과 비교해도 손색이 없을 정도로 이 연구 분야를 선도하는 그룹들이 많다. 이 분야에서 노벨상이 나오면 일본인이 나올 가능성도 높다고 보고 있다. 반면 이 학회에 참여한 한국인은 세 그룹 정도 밖에 없었다. 카이스트 오병하 교수님이 거의 유일하게 이 학계에서 한국인으로서 구조 생물학으로 크게 기여하시면서 한국의 존재를 나타내셨다. 이 학회의 chair인 Hirano는 본인에게 나중에 한국하고 같이 이 분야를 키우고 싶다는 말씀을 하셨다. 하지만, 이렇게까지 한국이 일본에 비해 이 분야에 대한 기여가 낮을 줄은 몰랐다. 그러면서 근본 원인이 무엇인가에 대해서도 질문을 하게 되었던 것 같다. 아직 한국의 연구분야는 누군가 한국인 대부라고 부를 수 있는 과학자가 있으면 그 분야는 강하게 되는 것 같은데 그런 인물이 없는 분야에 대해서는 약한 것이 아닌가 하는 생각이 들었다. 반면 일본은 이 분야에 대해 수십 년간 깊은 투자가 있었고, 또 이 분야 하나에 목 매달고 수십 년간 개척을 한 분들이 많다는 것도 새삼 느끼게 되었다. 그러면서 그들끼리 서로 협력 관계로 이 분야를 발전 시켰던 것이다. 이런 것을 보면서 한국의 과학은 어떤 방식으로 더 발전을 해야 할까에 대한 질문도 많이 던진 학회였던 것 같다. 이 분야를 보면 일본 한 나라가 유럽 전체 국가에서 이뤄지는 연구, 미국에서 이뤄지는 연구와 전혀 뒤쳐지지 않고 있다는 것을 더 느끼게 된다. SMC 단백질 연구의 시작은 일본인 과학자들의 이 분야에 뛰어듦과 동시에 시작되었다고 하더라도 과언이 아닌 듯 했다.
이 학회의 중요한 세션은 Condensin, cohesin, SMC5/6 복합체의 분자 기작에 대한 연구발표가 주를 이뤘고, 특별히 각각 이 단백질들의 어떤 구조 변화가 chromosome organization과 segregation을 일으키게 하는지 어떤 regulatory factor들이 각각 어떻게 작용하는지, 그리고 지금 hot한 어떻게 이 두 단백질들이 DNA loop extrusion을 하는지에 대한 주 talk이 이뤄졌다. 그리고는 이 단백질들 중 특별히 cohesin이 어떻게 암과 같은 질병과 연관이 있는지에 대해 깊이 다뤄졌다.
Ⅱ. 주요 발표 내용
1. Keynote session
이번 학회의 Keynote session은 영국의 Edinburgh 대학의 William Earnshaw가 condensin 단백질에 대해 발표를 했다. 이 condensin 단백질은 과거에는 non-histone scaffold라고 알려져 있었다. 또한, 염색체가 어떻게 organization 되는지를 이해하고자 하면서, Histone과 그 외의 단백질 기능에 대한 많은 debate가 있었다. Hirano는 condensin이 기능을 하는 것이 더 크다고 생각을 하는데, 그것이 이제는 옳았다고 William은 이야기 했다. 최근 Hirano는 histone 없이도 condensin I과 condensin II가 chromosome을 organization 할 수 있다고 Science지에 보고를 했다. 이처럼 chromosome organization 연구에 SMC 단백질이 차지하는 비중이 커졌다.
2. Prokaryotic SMC session
기본적으로 Eukaryotic cell과 Prokaryotic cell의 chromosome organization 되는 방식은 다르다. Harvard Medical School의 David Z. Rudner는 Bacillus subtillis의 박테리아에 있는 SMC complexes들이 chromosome arms를 origin에서 terminus까지 여행하듯이 움직인다고 했다. 이는 기존의 SMC 단백질들의 localization에 대해 첫 단추를 제공한 것이기도 하다. 이것으로 SMC 단백질이 DNA loop을 점차적으로 더 크게 만든다고 주장하였다.
KAIST의 오병하 교수님은 SMC의 구조와 구조 변화를 이야기를 했는데, prokaryotic의 SMC dimer의 전체 구조를 부분으로 나눠서 crystal 구조를 최초로 풀어냈고, 그것으로 전체를 재구성하는 방식으로 50 nm 길이의 SMC 단백질 dimer를 재구성 해냈다. 또한 crosslinking 방식으로 풀어진 구조가 합당함을 보였다. 또한 SMC 이합체의 구조를 Negative staining으로 측정을 했을 때에도 같은 I 형 구조를 가짐을 보였다. 질의 응답 시간에서 이 구조가 여전히 아직 이 분야에서는 핫한 이슈라는 것을 다시 느낄 수 있었다. 로잔 대학교의 Stephan Gruber는 SMC-ScpAB의 DNA chamber를 이야기 또한 아주 흥미로웠다.
체코의 Masaryk 대학의 Jan Palecek의 talk을 통해서 SMC protein의 non-SMC subunits을 구분 지을 때 KITE (Kleisin-Interacting Tandem winged-helix Element)와 HAWK (HEAT Associated With Kleisin)가 있다는 사실을 처음 알게 되었다. KITE는 prokaryotic SMC/ScpAB와 eukaryotic의 SMC5/6 complexes에 있다고 했다. 반면, HAWK의 경우 condensin과 cohesin에 있다. 이 두 단백질들의 구조 차이는 다른 조절 기능이 있을 것이라 여겨진다고 했다. 아직까지 서로 다른 모델들이 있는데 그것이 species마다 다른 것인지, 조건마다 다른 것인지 의문이 들었다. 또한 이 SMC 단백질들에 universality가 있는지도 궁금증을 갖게 되었다.
3. Chromosome biophysics and modeling
이 분야에 최근 Biophysics와 Simulation 쪽이 각광을 받고 있었다. 그 동안은 cell biology 쪽의 연구가 대세를 이루었으나, 단백질이 정제가 어느 정도 가능하고, 부분 부분 단백질 구조가 밝혀지면서, biophysics가 해결해야 하는 부분이 더 구체화 되었다. 나 역시 biophysicist로서 이 세션에서 발표를 할 수 있게 되었다.
Leonid Mirny (MIT, USA)는 SMC 복합체들은 interphase에서의 chromosome 응축과 metaphase에서의 segregation에서 loop extrusion을 할 것이라고 제안했다. 그리고 이 SMC 복합체들의 이런 loop extrusion 기능이 universal mechanism이라고 생각을 했다. Mirny는 cohesin 의 central role이 loop extrusion인데, CTCF는 loop extrusion을 조절하는 기능을 한다는 것을 밝혔고, 이것이 domain formation을 지시하는 기능을 한다고 했다. 이 CTCF가 없을 때에는 chromosomal domain이 없어짐을 발견했다. 또한 genome의 compartmentation이 cohesin과 CTCF의 loss에 따라 거의 영향을 받지 않았는데 이것의 원인은 아직 모른다. 또 재미있는 결과는 최근에 Nature에 accept되었던 single-nucleus Hi-C 기술로 chromosome structure를 single cell level에서 관찰했는데 이 결과 condensin I 또는 II의 서로 다른 loop formation rate를 관찰했고, 이것을 통해서 3D 모델을 만들어 냈다. 그의 시뮬레이션 결과는 정말 신기했다. Cohesin이 DNA를 loop extrusion을 한다는 것을 이용하여 아주 복잡한 두 DNA가 two sister chromatid가 되는 것도 보였다. 이는 이제까지 있는 cohesin이 loop extrusion을 만들 것이라는 가장 강력한 근거가 되는 것이다. 그의 발표는 이번 학회에서 가장 인기 있었던 발표 중 하나였지 않나 생각이 든다.

Excursion 시간에 신사에서 그림을 그리고 있는 Leonid Mirny (MIT, USA).
다음 학회의 포스터는 Mirny의 그림으로 만들자고 할 정도로 그림을 잘 그렸다. 그래서 논문을 쓸 때 연구실 사람들이 교수가 요구하는 그림 퀄러티 때문에 많은 스트레스를 받는다고 했다.
John F. Marko (Northwestern Univ., USA)는 단분자 생물물리 기법을 condensin에 적용한 결과를 발표를 했다. Condensin이 어떻게 DNA를 compaction하는지를 크게 보면 loop extrusion과 non-extrusion 모델을 생각할 수 있고, 또 다른 면에서는 하나의 condensin이 loop을 만들어 내는지 여러 개의 condensin 단백질이 함께 loop를 만들어내는지로 구분이 된다. John Marko의 경우에는 단분자 자기 집게법으로 condensin을 정제해 낼 때 gel-filtration에서 multi-complex와 single complex를 분리시켜서 ATPase activity를 재고, DNA의 compaction되는 정도, DNA와 binding되는 정도를 재었는데 multi-complex가 single complex랑 비교했을 때 다른 면들은 모두 비슷하나, DNA compaction되는 정도는 2배 정도 다르다는 것을 보였다. 이 때, 좀 debate가 생겼다. 아직 field에는 multi-complex model을 믿는 사람들이 있고, 그러지 않는 사람들이 있다. Multi-complex가 더 active하다는 것이 story를 만드는데 강한 증거가 될 수 있지만, 아직 multi-complex의 구조를 보여준 경우는 없기에 더 증거가 필요한 상황이라는 지적이 있었다.
4. Sister Chromatid cohesion
Douglas Koshland (UC Berkeley, USA)는 cohesin의 긴 coiled coil 부분의 새롭게 알려진 기작에 대해 설명을 하였다. 이 전까지 사람들은 이 coiled coil 부분이 단순하게 hinge와 head를 연결해 주는 connector 정도로 생각했고, 이것을 통해서 두 DNA 부분이 옭아맬 수 있다고 생각을 했다. 또한 최근 모델은 이 부분이 ring 모양에서 rod shape으로 변하는 과정 cohesin의 중요한 기작을 하게 만드는 원인이라고 생각을 했다. 그는 cohesin이 DNA tethering에 관여하고, DNA에 loading을 하는지, 안전하게 cohesin이 DNA에 binding하게 만드는 기작이 cohesin의 어떤 부분과 연관이 깊은지를 mutagenesis를 통해서 설명하였다.
Mitsuhiro Yanagida (OIST, Japan)는 일본의 SMC 단백질의 대부라고 볼 수 있는 과학자이다. 나이가 77세인데도 아주 활발하게 연구를 계속하고 있다. Yanagida가 말하면 이분의 업적 때문에 쉽게 부정할 수 없다고 한다. Yanagida의 가장 큰 업적 중 하나가 cohesin의 clip model인데 이것은 어떻게 cohesin이 DNA에 붙고, 떨어지는지를 설명하는 모델이다. cohesin의 구조를 clip 형태의 구조가 되어서 DNA를 잡는다고 하는 것을 제안했다. 그는 SMC hinge와 head가 접근해서 활과 같은 coiled coil이 만들어지면서 DNA과 결합됨을 모델화했다.
영국 Oxford의 Kim Nasmyth 그룹의 Madhusudhan Srinivasan은 cohesin이 이 chromosome에 결합되어 있을 때 cohsein의 topology와 이것이 HAWKs에 의해 어떻게 regulation 되는지를 설명했다. DNA가 anaphase에서 separase에 의해 cohesin의 cleavage가 일어나면서 cohesin이 풀릴 때까지, cohesin은 DNA를 붙들고 있게 된다. 이 결합은 Scc2, Pds5와 Scc3라고 불리는 HAWK에 의해 조절이 된다. Cohesin은 interphase에서 condensin은 mitosis에서 DNA가 chromatid 같은 구조가 되도록 마는데, 이들은 DNA의 large loop를 점진적으로 만들어냄으로 chromosome을 형성하게 하는데, Srinivasan은 Chip-seq과 covalent bonding을 이용하여 cohesin ring을 cross-linking 시켰다. 이것으로 어떻게 cohesin이 chromosome과 association 되는지를 보여줬다. 그 결과 cohesin이 individual DNA를 cohesin ring에 통과시킴으로 cohesin ring이 열리지 않아도, DNA를 entrapment하게 하거나 DNA loop를 만든다고 제안했다.
Yasuto Murayama (Tokyo Institute of Technology, Japan)는 cohesin이 어떻게 DNA 두 개를 잡아서 replication된 chromatid 두 개를 연결시키는지를 설명하였다. minichromosome을 이용하여 cohesin이 하나의 minichromosome을 loading한 다음에 다른 minichromosome은 ssDNA로 loading하고 또 그 ssDNA가 polymerization되는 것을 제안하였다. 흥미있는 모델이었다. 또한 이렇게 replication과 연관이 지어지는 것도 이제 이 쪽 연구를 하는 나로서는 신기한 생각이었다.
Hongtao Yu (Univ.of Texas, USA)는 cohesin loading과 release에 대한 구조와 기능적인 기작을 설명하였다. Cohesin ring이 열고 닫히는 것은 이것이 chromosome에 loading되고 release되는데 중요한 기작인데, 이것을 조절하는 것이 loading factor인 Scc2-Scc4와 releasing factor인 Pds5와 Wapl이 있다. 그리고 cohesin을 stabilization시키는 sororin도 있고, protease인 separase도 있다. Cohesin의 release는 Smc3 ATPase head와 Scc1의 N 말단의 interface에서 일어나는 것으로 알고 있고, 이것을 exit gate라고 한다. Yu는 crystal structure로 Scc2, Pds5, Wapl과 cohesin의 sub-complex인 SA2-Scc1과 separase를 보여줬다. 이 구조들은 separase에 의한 cohesin cleavage의 기작과 조절 기능을 보여주고, Scc2에 의한 cohesin loading과 Pds5-Wapl의 의존된 cohesin release를 보여준다. 생화학 기법을 이용해서 Pds5와 Scc2가 Smc3 ATPase head와 Scc1과 붙는 것이 경쟁 관계임을 보였다. 또한 Pds5랑 Scc2가 open gate 구조를 안정화 시킴으로 cohesin dynamics를 촉진시키고, 이것은 ATPase cycle 때 일시적으로 떠나는 것을 보였다.
Benjamin Rowland (NKI, The Netherlands)는 cohesin release factor인 Wapl에 대해 연구했다. Cohesin 복합체는 3D genome을 CTCF랑 함께 DNA loop를 만들어낸다. Cohesin의 DNA release factor인 Wapl은 loop extension을 제약시키고, CTCF의 잘못 orientation되는 site에서 DNA loop이 만들어지는 것을 방해한다. Scc2/Scc4 복합체가 chromatin loop이 커지고, topologically associated domains (TAD)를 형성시킨다는 것을 보였다. 이것으로 점진적으로 loop이 커지는 것으로 cohesin이 chromosome을 형성시킨다고 제안했고, TAD는 polyclonal collections of loop이라고 제안할 수 있었다. 또한 그는 Scc2/Scc4와 Wapl의 balance가 chromosome을 올바로 구조를 만드는데 중요하다고 이야기를 했다.
Jan-Michael Peters (IMP, Austria)는 CTCF가 어떻게 cohesin의 역할을 조절하는지에 대한 연구로 권위자이다. 그는 cohesin의 위치가 어떻게 지정이 되고, 이것이 어떻게 chromatin의 architecture를 만드는데 조정을 하는지는 여전히 아직 해결되지 않은 숙제인데, Jan-Michael Peters는 CTCF가 cohesin의 위치를 조절시키고, 이 위치가 chromatin의 architecture를 결국 만들어 낸다는 것을 전반사 현미경 위에서 DNA를 길게 늘여서 거기에 형광이 달린 cohesin과 CTCF로 보여주었다. 재미있게 DNA 위에서 sliding하는 cohesin이 CTCF가 달린 DNA를 넘지 못한 것을 보여주었다. 하지만, 아직 왜 cohesin의 sliding이 ATP independent하게 나타나는지는 이해하지 못하고 있는 상태라고 했다.
5. Condensin
이번 학회에는 Condensin만 집중해서 발표하는 세션도 있었다.
Christian H. Haering (EMBL, Germany)는 condensin 연구에 다양한 단분자 생물물리 기법을 많이 적용했다. Condensin 단백질 연구에 가장 중요한 것 중에 하나가 어떻게 condensin이 어떻게 DNA에 결합을 하는지이다. Christian Haering은 Crystallography로 Ycg1과 Kleisin이 DNA를 정전기적 상호작용으로 안전벨트처럼 붙잡고 있음을 보여주었다. 흥미로운 것은 linear DNA와 이 Ycg1-Kleisin은 아주 약한 결합에너지를 가졌는데 Kd가 2 μM인데 DNA를 plasmid로 바꿨을 때 Kd 값은 10배 이상이 작아졌다. 즉 이 safety belt가 한번 결합이 되면 topological하게 DNA가 결합이 되어 있어야 함을 이야기 한다. 그리고 자기집게 법으로 topological한 DNA가 condensin하게 결합이 되면 이것은 높은 salt 농도로도 분해시키지 못하는 것을 발견하였다.
Frank Uhlmann (The Francis Crick Institute, UK)는 condensin에 의한 mitotic chromatin landscape을 리모델링하는 것에 대해 발표를 하였다. Condensin이 mitotic chromatin architecture를 만드는지는 아직 모르는 상태인데, Uhlmann은 Hi-C 기술을 이용해서 mitotic chromosome condensation을 연구했다. Uhlmann은 condensin이 long-range, intra-chromosomal DNA 상호작용을 일으켜서 chromosome을 응축시킨다는 결과를 보여줬다.
Toru Hirota (JFCR, Japan)는 live cell imaging 기술(quantitate microscopic analysis)를 이용하여 condensin II와 topoisomerase IIa가 어떻게 함께 작용을 하는지를 보여주었다. Topoisomerase를 inactivation 시키면서 Chromosome의 morphology를 관찰하면서 연구했다.
6. Chromosome organization in meiosis/zygotes
이 세션에서는 condensin과 cohesin이 어떻게 감수분열에 관여를 하는지에 대한 보고가 있었다. 특별히 Tomoya Kitajima의 발표는 정말 인상적이었다. Chromosome segregation 는 유산과도 연관이 있지만, 다운 신드롬과 같은 유전 병에도 연관이 있다. 이게 나이가 들면서 cohesin 발현양이 줄어들어서 일어나는 이유도 있지만, oocyte의 크기 때문에 일어난다는 것도 보여준 것이다. Micropipette을 이용해서 oocyte의 cytosol을 빨아들임으로 oocyte의 크기를 조절하여 chromosome segregation을 관찰한 방법에 많은 감탄이 되었다.

이번 학회 Excursion 은 일본의 과거 양잠업을 했던 곳과 오래된 신사와 체리 밭에서 체리 따먹는 코스를 돌고 왔다. 이 시간을 통해서 다른 연구 팀과 많은 교류를 할 수 있었다.
7. SMC and Disease
SMC 단백질은 암, 유전 질병과 밀접한 관계가 있다. 따라서 SMC 단백질의 돌연변이와 병과의 관계가 활발하게 연구가 되어 오고 있다.
Matt Deardorff (Children's Hospital of Philadelphia, USA)는 병원에서 일하고 있고, SMC 단백질과 유전 병과의 관련성에 대해 연구를 하고 있었다. Cornelia de Lange Syndrome이라고 불리는 증상과, Rett-like Epileptic이라는 현상을 cohesin의 유전과 같이 연구를 하고 있었다. 특별히 Cornelia de Lange Syndrome 현상은 지적장애도 있고 다양한 현상이 있지만, 얼굴 형태가 보통 사람들과 다르고 얼굴 형태를 보면 이 병이 있음을 알 수 있다. 재미있는 것이 이 유전병을 일으키는 뮤테이션들이 cohesin에 몰려 있었다. Matt가 이 뮤테이션들을 병원 환자들을 통해서 찾아냈고, 이 특징들을 연구를 하였다. 하지만 아직 cell biology와 In vitro로 왜 그런 현상들이 일어났는지는 전혀 연구하지 못하고 있다고 했다. 이제 이런 유전병의 구체적인 증상을 연구하기 시작할 단계가 곧 오게 될 것 같다.

야외에서 함께 바비큐 파티를 했다.
Ⅲ. 총평
단백질 하나를 아주 집중적으로 다루는 학회는 처음이었다. 이 단백질의 시작부터 지금 활발하게 진행되는 과정까지 볼 수 있는 좋은 기회였다. 또한 기조 연설에서는 chromosome organization과 segregation의 연구 역사까지 볼 수 있었고, 그 연구 기간이 100년 이상이 된 분야라는 것이 더 실감할 수 있었던 시간이었다. 점점 더 SMC 단백질의 중요성이 밝혀지면서 한 동안 잠잠했던 이 단백질 연구가 지금 아주 활발하게 진행이 되고 있다. 또한 단백질 정제 기술의 발달로, in vitro 실험이 더욱 활발해졌고, 지금은 많은 cell biology 실험과 biochemistry 실험이 주를 이루며 특별히 Hi-C나 Chip-seq 기술이 많은 연구를 이루고 있지만, 많은 사람들이 내가 하고 있는 단분자 생물물리 기법에 관심을 가졌다. 물론 자기집게 법과 단분자 형광 기법을 이용한 연구도 많이 소개 되었다. 여러 이 분야의 선도하는 분들과 이야기 하다가 다들 단분자 생물물리 기법에 관심을 가졌다. 이 학회를 통해서 내가 어떻게 이 학계에 기여를 할 수 있는지에 대해 명확한 방향성을 얻었다. 그리고 가장 중요한 과제들이 나의 연구와 직접적으로 관련이 있다는 것도 더 느낄 수 있었다. 생물학의 패턴을 보면 먼저는 어떤 특정 유전자가 발견이 되고, phenotype을 알려고 하는 연구와 cell biology 연구가 집중적으로 진행이 되다가 단백질 정제를 통해서 구조 생물학과 in vitro 실험이 되기 시작하면서 더 detail한 연구가 진행이 된다. 거기에 이제는 최근 개발된 단분자 생물물리 기법이 생물학 연구 분야의 후반부를 장식하는 것 같다.

이번 학회의 Organizer인 Tomoko Nishiyama (Nagoya Univ), Tatsuya Hirano (Riken), Katsuhiko Shirahige의 사진.
이 분들 덕분에 이번 학회는 성공적이었고, 특히 Hirano는 마지막쯤, 피곤을 호소했다.
이 세분들은 이 분야에 막대한 업적을 남긴 분들이기도 하다.
☞ 자세한 내용은 내용바로가기 또는 첨부파일을 이용하시기 바랍니다.