

기술동향
새로운 N2O 환원 미생물의 유전체학과 생태학
- 등록일2019-11-18
- 조회수6307
- 분류기술동향
-
자료발간일
2019-11-14
-
출처
생물학연구정보센터(BRIC)
- 원문링크
-
키워드
#N2O#아산화질소#미생물
- 첨부파일
 pdf_0003364.pdf
pdf_0003364.pdf
새로운 N2O 환원 미생물의 유전체학과 생태학
< 목 차 >
1. 미생물 자원과 N2O 흡수원: N2O 환원 미생물에 관하여
2. N2O 환원 미생물의 진화와 유전체학
3. N2O 환원 박테리아 개체군의 생리학
4. N2O 환원 미생물 군집의 생태학
5. 생태계 기능에 미치는 파급효과
6. 결론
-------------------------------------------------------------------------
< 요약문 >
아산화질소(N2O) 배출은 환경파괴의 주요 원인 중 하나이다. 최근, 이 N2O를 환원시키는 미 생물이 발견되었다. 따라서, 미생물이 온실가스 중 하나인 N2O를 무해한 이질소가스로 환원시 키는 능력을 가졌다는 점이 이목을 끌고 있다. N2O 환원 능력이 있는 탈질화, 비탈질화 미생물 의 다양성은 예상보다 더 방대하다. 그동안 간과하던 자연계의 clade II 그룹은 N2O 흡수원으 로써 작동할 수 있는 비탈질화 N2O 환원 미생물의 대부분을 포함한다. 본 리뷰에서는 최근 N2O 의 유전체학, 생리학, 생태학적 기저에서 이뤄진 진보적인 연구 성과와 N2O 배출 억제를 위한 일환으로써 이 연구의 중요성을 논의하고자 한다.
1. 미생물 자원과 N2O 흡수원: N2O 환원 미생물에 관하여
무기질소화합물은 생물지구화학 순환 내에서 미생물이 유도하는 산화 환원 반응을 통해 변 환된다. 미생물은 생태계 내 질소를 유지하고 손실하는 일련의 과정을 조절하고, 어떤 형태의 질소 화합물을 분해할 지 결정한다. N2O는 여러 질소 순환 과정에서 생성되는 물질로 주요 온실가스 중 하나이자, 성층권 오존층 파괴의 원인이며, 100년 간 이산화탄소의 300 배에 해당하는 온난화 현상 을 일으켰다. N2O 배출량의 3분의 1을 차지하는 수생태계를 제외하고, 농토, 농업 용수, 가축 폐기물 등 토양생태계는 N2O 배출량의 대부분을 차지하고 있다. 시간이 지날 수록 비료 사용량이 증가하고 토지 이용 변화에 따라 토양 유기물이 광물화되면서 N2O 배출량은 점차 더 증가할 것으로 보인다.
생물권 안에 많은 N2O 공급원이 있지만 N2O 흡수원은 단 하나만 존재한다. 미생물 효소인 N2O 환원 효소(NosZ)가 N2O를 N2로 환원시킨다. 이 효소는 박테리아와 고세균에서 생성되어 N2O 를 N2로 환원시키는 완전한 탈질화(denitrification)를 일으키는 것으로 알려져 있다. Wolinella succino genes 와 Campylobacter fetus 를 포함한 몇몇 비탈질화 박테리아는 N2O를 유일한 전자수용체 로 이용하여 성장하는 것이 일찍이 보고된 바 있다. 최근 많은 미생물 분류군의 유전체(genome)가 구축되고 분석되면서, 기존의 예상보다 N2O 환원에 관여하는 두 탈질화, 비탈질화 미생물의 다양성 이 훨씬 크다는 것을 밝혀냈다. 탈질화 미생물과 대조적으로, 비탈질화 N2O 환원 박테리아와 고세균 은 N2O를 생성하기 보다 N2O 흡수원이 될 수 있는 가능성이 높다. 따라서 N2O 배출에 관하여 기 후변화 완화 전략을 개발할 때 농업생산과 오수처리시설 문제는 물론, N2O 환원 미생물의 능력은 더 욱 중요하게 다뤄져야 한다.
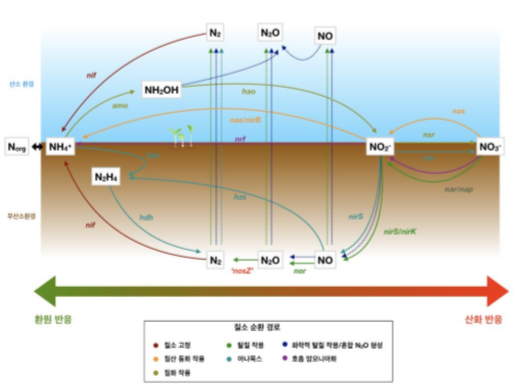
그림 1. 미생물의 질소 순환 경로
NosZ 단백질 계통도(phylogeny) 분석 결과, NosZ는 clade I 과 clade II 두 그룹으로 나뉘어지 고 있다. 이 clade들은 typical NosZ와 atypical NosZ로도 구분하지만, 본 리뷰에서는 clade I과 clade II NosZ로 구분하려 한다. Clade I 탈질화 미생물이 환원 반응하는 속도가 종마다 서로 다르기 때문에, 군집의 구성에 따라 N2O 흐름이 달라질 수 있는 것으로 보인다. Clade II에 대해 알려진 건 많지 않 지만, 점점 유전학, 생리학적 차이가 밝혀지고 있다. 본 리뷰에서는 N2O 환원 미생물들을 연구한 최 근 문헌들을 참조하여 유전학, 생리학, 생태학적 정보 관점에서 N2O 환원 미생물의 두 clade 간 또 는 clade 내 분서(niche differentiation)를 설명하고자 한다. 또한 이러한 연구가 토양을 비롯해 여러 N2O 배출 완화에 중요한 이유를 논의하고자 한다.
2. N2O 환원 미생물의 진화와 유전체학
N2O 환원 능력은 계통학적으로 널리 퍼진 형질(trait) 중 하나이다. N2O 환원 촉매소단위체를 부호화(Encoding) 하는 nosZ 유전자의 상동 유전자(homolog)들은 다양한 환경에 서식하는 열 두 종 의 고세균, 박테리아 유전체의 약 12%를 차지한다. 분석을 통해서 nosZ 계통도는 생물계통도와 일 치하며 수평 유전자 전달(horizontal gene transfer) 케이스를 제외하고 NosZ의 분화가 주로 수직 유 전 과정 중 발생한 것을 밝혀냈다. 이는 N2O 환원이 고대의 형질일 가능성을 보여주고 있다. 한편, 현재로부터 28~25억 년 전 시생대 후기 ‘산소 급증 사건(GOE; great oxygenation event)’ 전과 후 중 어느 시기에 N2O 환원 효소가 생성 되었는지 역시 의문이다. 동위원소 증거(isotopic evidence)에 따 르면 비탈질화 경로는 GOE 바로 뒤에 일어난 것으로 보인다. 효소의 주요 공동인자(cofactor) 중 하 나인 구리는 GOE 전 환경에서 낮은 농도는 아니지만 생물학적으로 이용 가능한 수준은 아니었다. 한편, N2O의 전구체인 NO는 GOE 전에도 있었고 철과 헴(heme)을 이용해 산소가 부족하더라도 N2O를 생성했을 것으로 예상된다. 또한 철(II)과 아질산염(nitrite)의 화학적 탈질작용 (chemodenitrification)을 통해 N2O가 생성될 수 있었다. 따라서, 아질산염 감소 시기에 일어난 N2O 의 축적과 동시에 N2O 환원 효소도 진화했을 것이다.
하지만 nosZ 유전자는 미생물 분류군을 통틀어서 볼 때 고르지 못한 분포를 보인다. 이는 근연종 간에도 그렇다. 이와 같은 다계통적 분포를 바탕으로, nos 유전자의 획득과 손실이 환경 요인 에 의해 결정된다는 가설을 세울 수 있다. 박테리아에서 종종 기능 상실(loss of function)은 곧 적합 성(fitness)을 증가시키기 때문에 결손(deletion)에 의한 유전자 손실은 공통적인 진화 현상 중 하나이 다. 실제로, 완전한 탈질화 미생물들은 질산염이 고갈되고 다른 자원이 제한적일 때 nosZ 발현을 하 향조절한다. 이는 nosZ 가 없는 경우 탈질과 관련된 아질산염 환원 효소 유전자가 유전체의 40% 가 량을 차지하는 이유이다. 이러한 현상은 무작위한 패턴이 아니다. 예를 들어, nosZ 는 곰팡이 유전체 에도 없고, 방선균류(Actinobacteria) 또는 엑시도박테리아류(Acidobacteria)의 유전체에도 존재하지 않는다. 이 분류군은 주로 낮은 자원가용성과 낮은 ph 조건의 토양 생태계와 관련이 있다.
nosZ 유전자는 두 개의 다른 N2O 환원 효소 그룹 clade I 과 clade II 그룹으로 구분된다. 이 에 대해 NosZ 단백질 계통도 결과 뿐만 아니라, nos 유전자 클러스터(NGC; nos gene cluster)의 구성, NosZ 전좌 경로(translocation pathway), 탈질화 또는 호흡 암모니아화(respiratory ammonification)와 관련된 다른 유전자와의 동시 발생 빈도의 차이가 같은 경향을 보인다. Clade I은 알파-, 베타-, 감마 프로테오박테리아로 구성되어 있다. 호염성 유리고세균(Haloarcula와 Natronomonas)의 nosZ 유전자 도 진화적 거리에 따라 clade I으로 분류된다. 신호 펩티드(signal peptide) 연구는 단백질이 세포질에 서 먼저 접힌 다음 세포질막을 가로질러 운반되는 twin-arginine 전좌 경로를 이용해 Clade I의 NosZ가 주변세포질로 전좌되는 것을 밝혀냈다. Clade I nosZ 을 포함한 NGC를 가진 개체는 단백질 조립과 구리 수송에 관여하는 DFYL 유전자 다음에 nosZ 유전자를 갖고 있다. 또한 NosZ로 전자 수 송을 담당하는 막결합 Fe-S 플라보단백질 유전자 nosR 의 존재에 따라 clade가 구분될 수 있다. 탈질 화 경로에서 다른 유전자와 nosZ 의 동시 발생을 분석했을 때, clade I nosZ 을 가진 개체의 83%는 두 아질산염 환원 효소 NirS 또는 NirK의 유전자를 가지고, 덕분에 clade I nosZ 를 지닌 개체는 더욱 완 전한 탈질화를 수행할 수 있다.
다양한 고세균과 박테리아 분류군은 clade II nosZ 를 가진다. Clade I과 다르게 clade II nosZ 를 지닌 개체들의 51%는 비탈질화 N2O 환원자들이다. 흥미로운 점은 이전 유전체 연구들에서 clade II nosZ 를 가진 개체의 약 27%는 호흡 암모니아화 생물이 주로 가지는 포름산염 의존성 아질산염 환 원 유전자 nrfA 를 가진다는 점이다. 하지만, Photobacterium profundum 과 Shewanella loihica 는 clade I NosZ를 가진 것을 보아 호흡 암모니아화가 clade II nosZ 가 있을 때만 일어나진 않는다. Shewanella loihica 는 몇몇 다른 박테리아( Opititus terrae 와 Bacillus spp .)들이 가지는 완벽한 탈질화와 호흡 암모 니아화를 수행하는 생물이다. 고세균과 호열성 박테리아를 제외하고 clade II nosZ 대부분이 Sec 전좌 경로와 관련된 신호 단백질을 가지고 있다. 이 전좌 경로는 단백질 접힘과 보조인자 결합이 일어나 고 있는 주변세포질로 미완성 폴리펩티드를 내보낸다. 두 clade 간 전좌 경로의 차이가 전좌 이외에 어떤 기능적 의미가 있는지, 또는 단지 nosZ 계통발생에 걸쳐 분류군의 분포와 일치하게 된 것인지 는 분명하지 않다. 그러나 , W. succinogenes , Bacillus azotoformans , Solitalea canadensis 를 포함한 clade II 개체의 연구들에서 알 수 있다시피 NosZ는 세포질막과 밀접한 관련이 있다. 이는 nosZ 바 로 뒤에 전자 수송에 관련된 막관통 단백질 유전자 nosB 가 있는 것을 통해 알 수 있다. 이러한 패 턴의 예외로는 clade I에 속하는 호염성 고세균이 nosB 를 포함한 NGC를 가진 것이 있다. 전체적으 로 유전체 분석은 clade I과 clade II nosZ 를 가진 개체 간의 차이를 명백하게 보여준다. 하지만 어떻 게 NGC 배열의 차이와 전좌 경로의 차이가 생리학적 차이를 이끌어내고, 더 나아가 분서를 일으키 는지 더욱 자세히 알기 위해서는 생화학적, 생리학적 연구들이 뒷받침되어야 한다.
...................(계속)
☞ 자세한 내용은 내용바로가기 또는 첨부파일을 이용하시기 바랍니다.