

기술동향
탄소 이화 작용 억제 현상(CCR): 영양소를 최대한으로 활용하는 다양한 방법
- 등록일2020-11-13
- 조회수4926
- 분류기술동향
-
자료발간일
2020-11-05
-
출처
생물학연구정보센터(BRIC)
- 원문링크
-
키워드
#탄소#이화작용#억제#CCR
- 첨부파일
 pdf_0003633.pdf
pdf_0003633.pdf
[BRIC View 리뷰논문요약]
탄소 이화 작용 억제 현상(CCR): 영양소를 최대한으로 활용하는 다양한 방법
우성화 (POSTECH)
요약문
대부분의 박테리아는 서로 다른 탄소원으로 구성된 혼합물로부터 기질을 선택적으로 사용할 수 있다. 선호 탄소원이 존재할 때, 비선호 탄소원의 이화 작용에 관련된 시스템의 발현, 때로는 효소 활성이 저해된다. ‘탄소 이화 작용 억제 현상(Carbon Catabolite Repression, CCR)’이라 불리는 이러한 현상은 전사 활성화, 전사 억제, 혹은 RNA 결합 단백질에 의한 번역 억제를 포함한 서로 다른 제어 기작을 통해 서로 다른 박테리아에서 수행된다. 또한, 많은 병원성 박테리아에서의 독성 인자 발현이 CCR에 의해 조절된다고 알려져 있다. 본 리뷰에서는 탄소원을 계층적으로 활용하기 위해서 여러 박테리아가 어떤 방식으로 진화되어 왔는지, 그 기작에 대해 다룰 것이다.
Key Words: Carbon Catabolite Repression (CCR), Carbon source
※ 본 자료는 Carbon catabolite repression in bacteria: many ways to make the most out of nutrients. Nat. Rev. Microbiol. 6, 613–624 (2008).의 논문을 한글로 번역, 요약한 자료입니다.
1. 서론
2. CCR에 대한 통괄 제어 기작
2.1. E. coli (대장균)에서의 기작
2.2. B. subtilis (고초균)에서의 기작
3. CCR에 대한 오페론 특이적인 제어 기작
3.1. 탄소원 운반 조절을 통한 제어
3.2. 전사 인자 조절을 통한 제어
4. 다른 박테리아에서의 CCR
4.1. Firmicutes (후벽균)에서의 기작
4.2. Actinobacter (방선균)에서의 기작
4.3. Pseudomonas putida와 Acinetobacter baylyi에서의 기작
5. 결론
1. 서론
대부분의 박테리아는 다양한 화합물을 탄소원으로 사용하는 것이 가능하다. 박테리아는 이러한 다양한 탄소원을 동시에 대사할 수도 있지만, 접근이 쉽거나 빠르게 자랄 수 있도록 하는 탄소원을 우선적으로 대사하기도 한다. 1942년 Jacques Monod에 의해 밝혀진, 대장균에서의 포도당(glucose)과 젖당(lactose)에 대한 선택적 탄소원 사용이 이에 대한 하나의 예시이다. 박테리아나 고등 생물을 대상으로 한 후속 연구를 통해 이런 선택적 탄소원 사용은 굉장히 흔한 현상이며, 많은경우에서 포도당이 선호 탄소원으로 취급됨이 확인되었다. 게다가, 포도당의 존재는 다른 비선호 탄소원의 대사를 억제했다. 포도당이 다른 탄소원에 비해서 선호되는 이와 같은 현상은 ‘포도당 억제(glucose repression)’ 또는 ‘탄소 이화 작용 억제 현상(Carbon Catabolite Repression, CCR)’으로 불린다. 오늘날 우리는 CCR을 ‘선호 탄소원이 존재할 때, 비선호 탄소원의 대사와 관련된 효소의 발현이나 활성이 억제되는 제어 현상’이라고 정의한다.
박테리아가 가지고 있는 5-10%의 유전자가 CCR에 의해 발현이 조절될 정도로, CCR은 대부분의 박테리아에서 굉장히 중요한 제어 현상 중 하나이다. 선호 탄소원을 선택적으로 활용하는 것은성장 속도를 결정한다. 따라서 다른 미생물과의 경쟁에서 살아남도록 하는 중요한 요인이기 때문에,이 CCR은 자연 환경에서의 생존 경쟁에 중요한 역할을 한다. 게다가, CCR은 병원성 유전자의 발현에도 중요한 역할을 하는데, 이는 영양 공급원이 없어진 박테리아가 쉽게 새로운 영양 공급원으로 접근하도록 하기 위한 하나의 기작이다. 따라서, 자연계 또는 병원성 박테리아에서 CCR이 진화되도록 한 주요한 추진력 중 하나는 더 빠른 성장을 가능하도록 하는 특정 탄소원의 선택적 활용 능력이라고 할 수 있다.
자연계에 존재하는 대부분의 종속 영양(heterotrophic) 박테리아와 더불어, 기능적 독립 영양(facultative autotrophic) 박테리아에서도 CCR은 관찰된다(기능적 독립 영양 박테리아에서는 유기탄소원이 존재할 때, 이산화탄소 고정과 관련된 유전자의 발현이 억제된다). 하지만, 예외가 몇 가지있다. Chlamydia trachomatis와 Mycoplasma pneumonia와 같이 영양이 풍부한 숙주 환경에 고도로적응된 몇몇 병원성 박테리아의 경우에는 CCR이 없는 것으로 보인다. 이러한 미생물은 몇몇 서식환경에만 적응되어 작은 유전체를 가지며, 대부분의 제어 기작이 없는 것으로 생각된다. 다른 예시로, Corynebacterium glutamicum에서는 포도당과 다른 탄소원이 동시에 대사되는데, 이는 굉장히 드문 현상이다. 마지막으로, Streptococcus thermophilus, Bifidobacterium longum, Pseudomonas aeruginosa와 같은 몇몇 박테리아에서는 포도당이 비선호 탄소원이고, 다른 선호 탄소원에 의해 대사 관련 유전자의 발현이 억제된다.‘Reverse CCR’이라고 불리는 이 현상은 해당 리뷰에서 다루지 않겠다.
많은 개체에서 대사 관련 유전자에 대한 CCR은 통괄 제어 기작과 오페론 특이적 제어 기작이 결합되어 수행된다. 해당 리뷰에서는, 이러한 기작을 전반적으로 설명하고, 다른 종류의 박테리아에서의 CCR에 대해서도 다루도록 하겠다.
2. CCR에 대한 통괄 제어 기작
CCR은 모델 미생물인 E. coli (대장균)과 B. subtilis (고초균)에서 가장 집중적으로 연구가 이루어졌다. 두 종 모두에서 CCR은 통괄 제어 기작과 오페론 특이적 제어 기작에 의해 작동된다. E. coli와 B. subtilis에서, 통괄 제어 기작의 결과는 비슷하여, 포도당이나 다른 선호 탄소원이 있을 때 비선호 탄소원대사를 가능하도록 하는 유전자가 발현되지 않는다. 하지만, 이러한 결과를 위한 분자적 기작은 완전히다르다. E. coli에서는 포도당이 있을 때 대사 유전자의 전사 활성화가 방해되며 CCR이 일어나는 반면, B. subtilis에서는 포도당이 있을 때 억제 단백질이 직접적으로 발현을 막는 방식으로 CCR이 일어난다. 이 두미생물에서 CCR의 기작이 서로 다르긴 하지만, CCR을 야기하는 신호 전달 경로에서 PEP(phosphoenolpyruvate)와 PTS (phosphotransferase system)이 가장 중요한 역할을 하는 것은 동일하다.
PTS는 기본적으로, 관련 효소들의 연속적인 인산화를 통해 탄소원을 운반하는 동시에 인산화시키는 시스템이다. 이러한 운반 시스템은 박테리아에만 존재한다. PTS는 개별적으로 발현되는 세 가지 단백질(enzyme I (EI), Histidine protein (HPr), enzyme II (EII))로 이루어져 있다. EI는 PEP로부터 인산기를 넘겨 받아 연속적인 인산화의 시작을 하고, HPr의 His15 부분을 인산화시키는 역할을 한다. 그 다음으로 인산화된HPr은 기질 특이적인 수송 단백질 EII의 A 도메인에 인산기를 전달한다. 이후, B, C 도메인을 통과하는 특정 기질로 인산기가 전달된다. PTS 단백질 사이의 인산화는 가역적이다. 따라서, PTS 단백질의 인산화된정도는 PTS에 의한 수송 효율과 해당 과정의 흐름량을 대표하는 PEP/피브르산 비율에 영향 받는다. 영향환경과 세포 대사 상태에 대한 반응에 따라 PTS 단백질의 인산화된 정도가 동적으로 변화하는 것을 통해서, PTS를 통한 신호 전달 및 유전자 발현 제어가 가능해진다. 장내세균과에서의 CCR은 대부분 EIIA에 의해 조절되는 반면에, 후벽균과에서의 CCR은 대부분 HPr에 의해 조절된다. 추가적으로 HPr (His-P)은 대사효소나 전사 인자의 인산화에 관여하여 그들의 활성을 조절하며, EI나 EII 또한 그 자체로 유전자 발현을조절하거나, 효소의 활성을 조절하기도한다. 흥미롭게도, 유전체 비교 분석(comparative genome analysis) 결과에 의하면, PTS는 원래 신호 전달 시스템이었고, 수송 기능은 이후의 진화 과정을 통해 획득된 것으로보여진다.
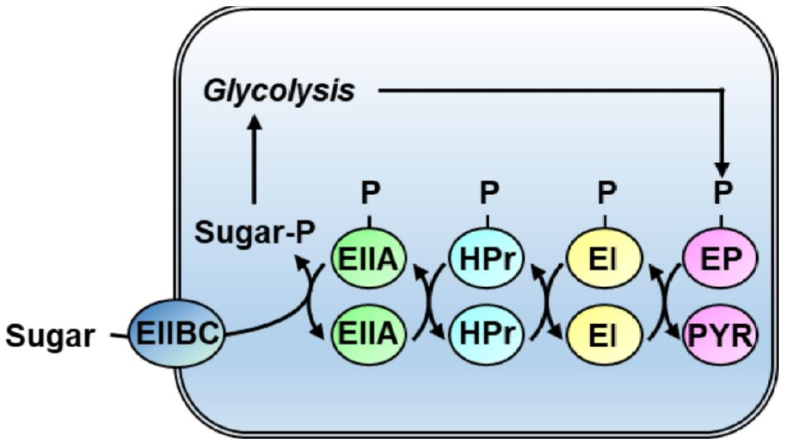
그림 1. PTS (phosphotransferase system)의 작용 기작.
...................(계속)
☞ 자세한 내용은 내용바로가기 또는 첨부파일을 이용하시기 바랍니다.
-
다음글
- 코로나19 백신 가격 전망 및 가격 결정 모델
-
이전글
- 감염대응 분자진단 및 바이오칩_바이오의료융합_기초연구본부 선정 R&D 이슈 연구동향(4) [R&D Brief 2020-25]
지식
- BioINwatch 기후변화 대응기술로 미생물 패러다임 확산 2025-07-08
- BioINpro [KRIBB 워킹그룹] 생물학적 CCU 기술 개요 및 주요 동향 2023-07-03
- BioINwatch 탄소나노튜브와 머신러닝을 활용한 혈액검사로 난소암을 조기에 진단할 수 있는 디지털바이오기술 2022-11-03
- BioINwatch MP(microbial protein) 20% 대체로 산림 벌채 및 CO2 배출 50% 감소 효과 2022-05-10
- BioINwatch 이산화탄소를 식량자원으로 전환하는 무세포 기반의 인공합성회로 개발 2021-12-02
동향